



Publicado
2004-01-01
Modificaciones técnicas en el uso de microsatélites y AFLP para el estudio poblacional de diversas especies de peces en el río Sinú, Colombia
Modified techniques used for microsatellite and AFLP for the population study of divers species at the Sinu river fish, Colombia
Palabras clave:
ADN, biología molecular, genotipificación, ictiología, DNA extraction, molecular biology, genotypification, ichthyology. (es)
En Colombia se han realizado pocos estudios de conservación genética aplicando técnicas de biología molecular. Debido a la complejidad del proceso de estandarización de las técnicas de laboratorio se presentan los protocolos y las modificaciones realizadas durante el estudio genético de cuatro especies ícticas del río Sinú, con el fin de que sean útiles como guía para futuros estudios, no sólo para peces sino para otras especies. Se realizaron modificaciones de los protocolos originales en la extracción de ADN, la cual fue llevada a cabo con un kit comercial, en las reacciones de PCR para la obtención de marcadores microsatélites y AFLP, así como en la genotipif icación de alelos. Las variaciones realizadas fueron distintas para cada una de las especies trabajadas.
Palabras clave: extracción ADN; biología molecular; genotipificación; ictiología; DNA extraction; molecular biology; genotypification; ichthyology.
Few genetic conservation studies carried out in Colombia have applied molecular biology techniques. Protocols and modifications arising from the genetic study of four fish species from the Sinú river basin are presented as guidelines for future work in this field (and with other species) as laboratory standardisation processes are complex. The original commercial kit's DNA extraction protocols were modified as were those for PCR reactions for obtaining micro-satellite markers and AFLPs, as well as allele genotypification. Particular variations were made f or each of the species studied.
Key words: DNA extraction; molecular biology; genotypification; ichthyology.
|
|
|||
|
Modificaciones técnicas en el uso de microsatélites y AFLP para el estudio poblacional de diversas especies de peces
en el río Sinú, Colombia
Modified techniques used for microsatellite and AFLP for the
population study of divers species
at the Sinu river fish, Colombia
Natalia Lamprea*, Liliana López*, Diana Santacruz*, Jimena Guerrero**, Consuelo Burbano***
|
|||
|
|
|||
|
RESUMEN
En Colombia se han realizado pocos estudios de conservación genética aplicando técnicas de biología molecular. Debido a la complejidad del proceso de estandarización de las técnicas de laboratorio se presentan los protocolos y las modificaciones realizadas durante el estudio genético de cuatro especies ícticas del río Sinú, con el fin de que sean útiles como guía para futuros estudios, no sólo para peces sino para otras especies. Se realizaron modificaciones de los protocolos originales en la extracción de ADN, la cual fue llevada a cabo con un kit comercial, en las reacciones de PCR para la obtención de marcadores microsatélites y AFLP, así como en la genotipif icación de alelos. Las variaciones realizadas fueron distintas para cada una de las especies trabajadas.
Palabras clave: extracción ADN, biología molecular, genotipificación, ictiología.
ABSTRACT
Few genetic conservation studies carried out in Colombia have applied molecular biology techniques. Protocols and modifications arising from the genetic study of four fish species from the Sinú river basin are presented as guidelines for future work in this field (and with other species) as laboratory standardisation processes are complex. The original commercial kit's DNA extraction protocols were modified as were those for PCR reactions for obtaining micro-satellite markers and AFLPs, as well as allele genotypification. Particular variations were made f or each of the species studied.
Key words: DNA extraction, molecular biology, genotypification, ichthyology.
|
|||
|
|
|||
|
INTRODUCCIÓN
El uso de técnicas moleculares ha permitido evaluar varios aspectos genéticos de las especies silvestres, algunas en peligro de extinción, como la variación genética de poblaciones disminuidas demográficamente, la existencia de flujo génico entre poblaciones, la estructura genética y la definición de unidades genéticas para conservación (Smith y Wayne, 1996).
|
Con este objetivo se han utilizado marcadores genéticos como el ADN mitocondrial, los loci isoenzimáticos (Queller et al., 1993), los microsatélites o repeticiones cortas en tándem (STR) (Estoup et al., 1998), los polimorfismos de amplificación al azar (RAPD), de longitud (AFLP) y de restricción (RFLP) (Vos et al., 1995).
|
||
|
|
|||
|
Biólogas. Universidad Nacional de Colombia. Correo electrónico: lamprea@telesat.com.co, diana_santacruz@yahoo.com, lililokl@yahoo.com
Bióloga Pontificia Universidad Javeriana, M. Sc. Genética. Correo electrónico: cguer@tutopia.com
Profesora asistente. Universidad Nacional de Colombia, M. Sc. Genética. Correo electrónico: cburbano@ciencias.unal.edu.co. Dirección postal: Departamento de Biología, Universidad Nacional de Colombia. Ciudad Universitaria. Bogotá. A.A. 14490.
Recibido: enero 22 de 2004. Aceptado: mayo 21 de 2004.
|
|||
|
|
|||
|
72
|
|||
|
|
|||
|
|
|||
|
MODIFICACIONES TÉCNICAS EN EL USO DE MICROSATÉLITES Y AFLP
|
|||
|
|
|||
|
Los microsatélites son secuencias de repetición en tándem de 1 a 6 pares de bases. Han demostrado ser útiles dadas sus características como alto polimorfismo y amplia distribución en el genoma de todos los organismos. En general son selectivamente neutros y de relativo fácil manejo en el laboratorio (Goldstein y Pollock, 1997).
Los marcadores AFLP, por su parte, se basan en polimorfismos de longitud de fragmentos amplificados por medio de la reacción en cadena de la polimerasa (AFLP-PCR). Además de ser marcadores codominantes, no dependen de los estados alélicos ni de la expresión génica (Mueller y Wolfenbarger, 1999).
La aplicación de la biología molecular en especies silvestres poco trabajadas presenta varias complicaciones puesto que no existen protocolos estandarizados para trabajar tipo de muestras como músculo agalla alevino, etc.; no hay cebadores específicos diseñados para estas especies y es poco el intervalo de posibilidades para escogerlos; si las especies son escasas en medio silvestre no se pueden usar técnicas invasivas. Esto implica que la mayoría de estos estudios requieran un largo tiempo de estandarización en las técnicas de laboratorio. En Colombia han sido muy escasos los estudios de diversidad genética de especies endémicas, especialmente en peces (Bejarano, 2001; Castiblanco, 2003), por lo que es de gran importancia divulgar el procedimiento de estandarización ya que puede ser útil como protocolo de trabajo en futuros estudios de conservación genética para otras especies silvestres.
A continuación se presentan las modificaciones realizadas para la extracción de ADN a partir de tejido de Caquetaia krausii (mojarra amarilla), Brycon moorei sinuensis (dorada), Prochilodus magdalenae (bocachico) y Sorubim cuspicaudus (bagre blanco), usando un kit comercial. En cuanto a la aplicación de marcadores moleculares se presentan los protocolos utilizados para la obtención de marcadores microsatélites en C. krausii, B. moorei sinuensis y P. magdalenae por medio de amplificación cruzada, ya que aún no se han desarrollado cebadores específicos para estas especies, y para la obtención de marcadores AFLP en S. cuspicaudus.
|
MATERIALES Y MÉTODOS Extracción de ADN
Colección y preservación del tejido. La colección de las muestras se realizó en distintas localidades de la cuenca del río Sinú así como en estaciones piscícolas de la zona, tomando un trozo de tejido muscular o de branquia. En algunas de las estaciones piscícolas se colectaron alevinos. Todas las muestras se preservaron en etanol al 96%. A su llegada al laboratorio se realizó un recambio de etanol y se almacenaron a 4 °C.
Extracción. La extracción se realizó utilizando el kit (Wizard Genomics DNA Purification kit) de Promega®, el cual trabaja con tres soluciones principales: una solución SDS-hipotónica para lisis nuclear, RNAsa para degradar ARN y solución precipitadora de proteínas. El ADN es precipitado usando isopropanol (Promega, 1996). Se siguió el protocolo para tejido animal de hígado de ratón en las muestras de B. moorei sinuensis, P. magdalenae y S. cuspicaudus, mientras que para C. kraussii se siguió el protocolo para tejido de cola de ratón, debido a que éste reduce los residuos de proteína que se producen al usar el protocolo para tejido animal de hígado y que afectan la calidad del ADN en esta especie.
En general, el cambio más importante se realizó en el paso de maceración y en la cantidad de tejido empleado. Para las muestras de músculo se utilizaron aproximadamente 0.3 cm3 de tejido. Para el tejido de branquias se trabajó con cuatro filamentos de éstas y para los alevinos, con el tercio posterior del animal (tabla 1).
C. kraussii es la única especie que requiere un pretratamiento del tejido donde se adiciona EDTA y proteinasa K junto con la solución de lisis nuclear, en el primer paso del protocolo, al igual que se incrementa el tiempo de incubación de esta mezcla; mientras que el tejido muscular de S. cuspicaudus requiere maceración con nitrógeno líquido para disgregar el tejido (tabla 1).
La calidad del ADN se verificó mediante geles de agarosa al 0.8%, en cámara de electroforesis horizontal a 80V durante 30 minutos, con tinción de bromuro de etidio. Una banda nítida indica una ex-
|
||
|
|
|||
|
73
|
|||
|
|
|||
|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
REVISTA COLOMBIANA DE BIOTECNOLOGÍA VOL. VI No.1 Julio 2004 72 - 78
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
tracción positiva (figura 1). La eficiencia del proceso de extracción fue similar en las especies trabajadas (94 a 96%). Posteriormente se procedió a realizar la amplificación de segmentos de ADN por medio de la reacción en cadena de la polimerasa (PCR).
|
Aplicación de marcadores microsatélites
Amplificación en Caquetaia kraussii. Se probaron 12 parejas de cebadores de la especie Oreochromis niloticus, correspondientes a los loci: UNH106, UNH109,
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
74
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
MODIFICACIONES TÉCNICAS EN EL USO DE MICROSATÉLITES Y AFLP
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
UNH123, UNH129, UNH132, UNH174,UNH189, UNH191, UNH192, UNH209, UNH216, UNH231, seleccionados por poseer regiones repetitivas de dinucleótidos y temperaturas de alineamiento altas para la amplificación en O. Niloticus (Kocher, 1997).
|
1.20 min. de alineamiento a 47-54 °C, 1.20 min de extensión a 68 o 72 °C y extensión final a 68 o 72 °C por 3 min.
Los productos amplificados fueron almacenados a -20 °C. Las amplificaciones se verificaron por medio de electroforesis en geles de agarosa 3% teñidos con bromuro de etidio a 70 V por 50 minutos en cámara de electroforesis horizontal.
De las doce parejas de cebadores probadas, seis amplificaron positivamente en C. kraussii (tabla 2).
Amplificación en Prochilodus magdalenae y Brycon moorei sinuensis. Se utilizaron primers desarrollados para una especie relacionada, Piaractus mesopotamicus (Calcagnotto et al., 2001). Como control positivo se utilizó ADN de un individuo de la especie Piaractus brachypomus en la cual la eficacia de los primers fue evaluada por los mismos autores.
Cada reacción de PCR se realizó en un volumen total de 12.5 \iL con: 2.0 ^L de ADN, 10 mM de Buffer B de amplificación (100 mM Tris-HCl, 500 mM KCl, 1 % de Triton), 2.5mM de MgCl2 (excepto para el bocachico en el locus Pme20 donde se usaron 2.0 mM), 200 \iM de solución de dNTP, 0.8 ^M de solución de cada primer y 0.5 U de Taq polimerasa. Las condiciones de la reacción se realizaron de la siguiente manera: desnaturalización inicial del ADN a 95 °C por 4 min, 30 a 40 ciclos con 30 segundos de desnaturalización a 95 °C, 30 segundos de alineamiento a 44-57 °C y 30 segundos de extensión a 68-
72 °C. Por último, una extensión a 68-72 °C
por 10 min.
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Figura 1. Gel de Agarosa 0.8% donde se verifica la calidad y cantidad de ADN en C. kraussii. Flecha: banda de ADN.
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
La estandarización de la amplificación se inició siguiendo el protocolo original de Kocher y colaboradores (1998), utilizando como control positivo ADN de O. niloticus. Para lograr una amplificación exitosa en C. kraussii se realizaron cambios en la temperatura de alineamiento y la concentración de MgCl2 (tabla 2).
La PCR se realizó en un volumen final de 25 ^L, con: 10mM de Buffer B de amplificación (100mM Tris-HCl, 500mM KCl, 1% de Triton), 2.5-3.0 mM de MgCl2, 0.16 - 0.18 mM de dNTPs, 0.16 ^M de primer, 1.5 ^L de ADN y 0.5 U de Taq polimerasa. Las condiciones para la PCR fueron: 1 min inicial a 95 °C, seguido de 26 a 32 ciclos a 1 min de desnaturalización a 95 °C,
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Tabla 2. Loci amplificados en C. kraussii y condiciones específicas para PCR.
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
En el caso del P. magdalenae para los locus Pme5 y Pme14, fue necesario hacer los primeros tres ciclos con una temperatura de alineamiento baja: 45 °C para Pme5 y 44 °C para Pme14. Para el resto de ciclos (37) se usó una temperatura mayor (tabla 3).
Los productos amplificados fueron verificados con el mismo método que para C. kraussii. De los ocho cebadores desarrollados por Calcagnotto y colaboradores (2001) seis fueron exitosamente genotipificados para B. moorei sinuensis y cinco para P. magdalenae. Para el locus Pme28 en B. moorei sinuensis sólo se observaron bandas en regiones no correspon-
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Tm: temperatura de alineamiento; N°: número de ciclos para cada temperatura de alineamiento.
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
75
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
REVISTA COLOMBIANA DE BIOTECNOLOGÍA VOL. VI No.1 Julio 2004 72 - 78
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
cias de peso molecular de 50 bp y 10 bp en por lo menos un carril por gel, y la tinción se realizó en solución de bromuro de etidio de 5 mg/800 mL por 12 minutos (figura 3).
La asignación de alelos se realizó con el programa Gene Tools (Hitachi Genetic Systems®), el cual permite cuantificar el tamaño molecular de cada banda con respecto al marcador de peso molecular utilizado (Hitachi, 1998).
En general, el polimorfismo de los loci amplificados para C. kraussii
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
dientes a los microsatélites. De manera similar, para los loci Pme2 y Pme4 en P. magdalenae se obtuvo una efectividad de amplificación muy baja, 39 y 29%, respectivamente. Por esta razón, los loci anteriormente mencionados no fueron tomados en cuenta.
Genotipificación de microsatélites. Para las especies C. kraussii, B. moorei sinuensis y P. magdalenae, la genotipificación se realizó en geles denaturantes de poliacrilamida 6% (acrilamida/bisacrilamida 29:1, 4.6M urea, TBE 1X) en cámara de electroforesis vertical, con 123 V durante 6 horas. Se utilizaron referen-
|
fue alto, encontrándose entre 10 y 24 alelos por loci
con un tamaño similar al reportado para O. niloticus. La diferencia entre los alelos de mayor y menor tamaño estuvo entre 20 y 45 pares de bases (tabla 4).
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
En todos los casos el intervalo de tamaño de los loci microsatélites encontrados en el bocachico y la dorada fue mayor al obtenido por Calcagnotto y colaboradores (2001) para Piractus mesopotamicus, aunque en la mayoría de los casos los rangos se sobrelapan. Se encontró también un alto polimorfismo para todos los loci analizados con un número de alelos por locus entre 5 y 17 (tabla 5).
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
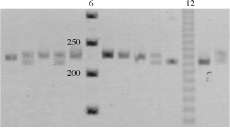
Figura 3. Gel denaturante de poliacrilamida 6% con tinción de bromuro de etidio para genotipificación de muestras de P. magdalenae para el locus Pme20. Carril 6: patrón de peso molecular de 50 bp. Carril 12: patrón de peso molecular de 10 bp. Las muestras se observan en un intervalo de 200 a 250 bp.
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
76
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
MODIFICACIONES TÉCNICAS EN EL USO DE MICROSATÉLITES Y AFLP
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
lección mediante pre-amp primer mix en 20 ^L el cual contenía: buffer 10 x, Taq polimerasa, MgCl2 y DNA ligado. Se realizaron 20 ciclos a 94 °C por 30 seg, 56 °C por 60 seg, 72 °C por 60 seg. Los productos se diluyeron 1:50 (ADN ligado: buffer TE) y luego se verificaron en geles de agarosa al 0.8%; las bandas que se observan en este punto son más intensas que el ADN extraído, con un barrido continuo pero más definido que las digestiones, indicando una preamplificación exitosa.
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Aplicación de marcadores AFLP en Sorubim cuspicaudus
La técnica de AFLP está basada en la amplificación de subgrupos de fragmentos de restricción genómicos usando PCR. El ADN se corta con enzimas de restricción y los adaptadores de doble cadena son ligados a la terminación de los fragmentos de ADN para generar el ADN molde utilizado en la amplificación. Las secuencias de los adaptadores y los sitios de restricción adyacentes sirven como lugares de unión para la posterior amplificación de los fragmentos de restricción (Vos et al., 1995).
Para genomas complejos, como el de S. cuspicaudus, es necesario hacer una preamplificación y una amplificación con el fin de reducir el número de fragmentos, facilitando la lectura en los geles de poliacrilamida (Vos et al., 1995).
El proceso de obtención de marcadores AFLP se realizó bajo el protocolo de AFLP Análisis System I y AFLP Starter primer kit de Life Technologies GIBCO BRL ® (GIBCO BRL, 1999).
Digestión y ligación. Para las digestiones se mezclaron 2.5 ^L de buffer 5X, 2.5 ^L de DNA, 1 ^L de EcoRI/MseI, 6.5 ^L de agua destilada y se realizó una incubación a 70 °C por 15 min, la cual fue visualizada en geles de agarosa al 0.8% teñidos con bromuro de etidio. Posteriormente se mezclaron 9 [j,L del producto de digestión con 8,5 ^L de EcoRI/ MseI, 1 [j,L de T4 ligasa y se incubó la mezcla a 20 °C por 2 horas. El paso de preamplificación se realizó con una dilución 1:10 de esta mezcla.
Preamplificación. Los productos de la ligación se preamplificaron con un primer de un nucleótido de se-
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Amplificaciones. A partir de las preamplificaciones se realizó una segunda amplificación con primers de tres nucleótidos de selección: 0.5 ^L de EcoRI (E-AGC) y 4.5 nL de MseI (M- CTG), 2.0 |iL buffer 10 x, 0.1 ^L Taq polimerasa con MgCl2, 7.9 ^L de agua destilada y 5 ^L de ADN digerido y preamplificado (dilución 1:50). La reacción se llevó a cabo en termociclador de la siguiente manera: 1 ciclo inicial a 94 °C de desnaturalización por 30 seg, 65 °C de alineamiento por 30 seg y 72 °C de extensión por 60 seg; posteriormente 12 ciclos bajando la temperatura de alineamiento 5 °C entre un ciclo y el siguiente. Por último, 23 ciclos a 94 °C de desnaturalización por 30 seg, 56 °C de extensión por 30 seg y 72 °C de extensión por 60 seg. El producto de la amplificación se evidenció en geles de agarosa al 1% teñidos con bromuro de etidio mediante un barrido continuo.
Genotipificación. La amplificación selectiva con tres nucleótidos de selección permitió obtener fragmentos más definidos en geles denaturantes de poliacrilamida al 6% (acrilamida:bisacrilamida 29:1, úrea 4,6M y TBE 1x).
Los geles de poliacrilamida al 6% se corrieron en cámara de electroforesis vertical de 40 cm por 20 cm a 1800 V por 1 hora y fueron revelados con tinción de plata (Promega Silver Staining Kit ®).
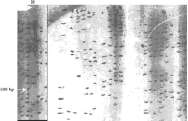
El tamaño aproximado de las bandas se estimó por comparación con un patrón de peso molecular de 100 pares de bases (pb). Este procedimiento permitió determinar 55 bandas polimórficas en un rango de 400 a 1000 pares de bases; las bandas de tamaño menor o mayor a este rango fueron consideradas no significativas (figura 2).
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
77
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|||
|
REVISTA COLOMBIANA DE BIOTECNOLOGÍA VOL. VI No.1 Julio 2004 72 - 78
|
|||
|
|
|||
 UOOtpM
|
AGRADECIMIENTOS
Las autoras agradecen a la empresa Urrá S.A. ESP por la financiación del proyecto Caracterización genética de cinco especies ícticas del río Sinú. Igualmente expresan su agradecimiento al Departamento de Biología de la Universidad Nacional por permitir el uso del laboratorio de conservación genética.
BIBLIOGRAFÍA
Bejarano P., 2001. Estructura genética de las poblaciones ex situ de Crocodrylus intermedius (Graves, 1819) en Colombia, mediante marcadores AFLP. Bogotá: Colombia: Departamento de Biología. Universidad Nacional de Colombia. 152.
Castiblanco J., 2003. Caracterización genética de dos poblaciones de Tilapia (Orechromis niloticus) en la cuenca del río Sinú. Bogotá, Colombia: Departamento de Biología. Universidad de los Andes. 180.
Calcagnotto, D.; Russello. M.; Desalle, R. 2001. Isolation and characterization of microsatellite loci in Piaractus mesopotamicus and their applicablility in other Serrasalminae Fish. Molecular Ecology Notes. 1: 245-247.
Estoup, A.; Rousset, F.; Michalakis, Y.; Cornuet, J. M.; Adriamanga, M.; Guyomard, R. 1998. Comparative analysis of microsatellite and allozyme markers: a case study investigating microgeographic differentiation in brown trout (Salmo trutta). Molecular Ecology. 7:339-353.
Gibco, BRL. 1999. AFLP Analysis system I & AFLP starter primer kit. Instruction Manual Life Technologies.
Goldstein, D. B.; Pollock, D. D. 1997. Launching microsatellites: a review of mutation processes and methods of phylogenetic inference. Journal of Heredity. 88:335-342.
Hitachi. 1998. Genetic systems. Gene tools users manual, Synoptics limited.
Kocher, T. 1997. CA-Tilapia.html. http://tilapia.unh.edu/ WWWPages/TGP/CA-Tilapia.html. Provee vínculos a la base de datos del GeneBank, donde se depositaron las secuencias de cada locus y los primers que se diseñaron para amplificarlos. Además, provee vínculos para información de loci aislados de Cíchlidos del lago Malawi. Revisado en marzo de 2003.
Kocher, T.; Lee, W.; Soboloewska, H.; Penman, D.; Mcandrew, B. 1998. A genetic linkage map of a cichlid fish, the tilapia (Oreochromis niloticus). Genetics. 148: 1225-1232.
Mueller, U.; Wolfenbarger, L. 1999. AFLP genotyping and fingerprinting. TREE. 14(10):389-394.
Promega. 1996. Protocols and applications guide. Third Edition.
Queller, C. R.; Strassman, I. E.; Hughes, C. R. 1993. Microsatellites and kindships. TREE. 8: 285-288.
Smith, T.; Wayne, R. 1996. Molecular Genetic Approaches in Conservation. New York: Oxford University Press Inc. 332.
Vos, P.; Hogers, R.; Bleeker, M.; Reijans, M.; Van, de Lee T.; Hornes, M.; Frijters, A.; Pot J.; Peleman, J.; Martin, K.; Zabeau, M. 1995. AFLP: a new technique for DNA fingerprinting. Nucleic Acid Research. 25(21): 4407-4414.
|
||
|
Figura 2. Gel denaturante de poliacrilamida (PAGE) al 6% con tinción de nitrato de plata revelando los productos amplificados de individuos de S. cuspicaudus. M: marcador de peso 1500-100 pb.
CONCLUSIONES
Se logró la estandarización para la extracción de DNA a partir de tejido muscular, de alevino y de branquia para las especies Caquetaia kraussii, Brycon moorei sinuensis, Prochilodus magdalenae y Sorubim cuspicaudus, utilizando un kit comercial (Wizard genomics de Promega®), con una efectividad promedio del 95%.
El protocolo de los procedimientos realizados en laboratorio fue distinto para cada tejido utilizado y para cada especie. Por esta razón se presenta un protocolo diferente en cada caso.
Con respecto a métodos de extracción clásicos, éste aumenta la eficiencia y representa una ventaja en tiempo de extracción y en reducción de desechos en comparación con métodos basados en solventes orgánicos.
El éxito de amplificación de los microsatélites fue variable para cada una de las especies evaluadas con estos marcadores, siendo de 50% (6 positivos de 12 cebadores probados) para C. kraussii, 75% (6 positivos de 8 cebadores probados) para B. moorei sinuensis y 63% para P. Magdalenae (5 positivos de 8 cebadores probados).
La importancia del presente estudio radica en que fue posible aplicar técnicas moleculares al estudio genético de poblaciones de cuatro especies ícticas del río Sinú, lo cual permitió no sólo el análisis del estado genético actual de las poblaciones estudiadas, sino que se logró establecer una guía de trabajo de laboratorio para su uso futuro en trabajos de conservación genética.
|
|||
|
|
|||
|
78
|
|||
|
|
|||
Cómo citar
APA
Lamprea, N., López, L., Santacruz, D., Guerrero, J. & Burbano, C. (2004). Modificaciones técnicas en el uso de microsatélites y AFLP para el estudio poblacional de diversas especies de peces en el río Sinú, Colombia. Revista Colombiana de Biotecnología, 6(1), 72–78. https://revistas.unal.edu.co/index.php/biotecnologia/article/view/584
ACM
[1]
Lamprea, N., López, L., Santacruz, D., Guerrero, J. y Burbano, C. 2004. Modificaciones técnicas en el uso de microsatélites y AFLP para el estudio poblacional de diversas especies de peces en el río Sinú, Colombia. Revista Colombiana de Biotecnología. 6, 1 (ene. 2004), 72–78.
ACS
(1)
Lamprea, N.; López, L.; Santacruz, D.; Guerrero, J.; Burbano, C. Modificaciones técnicas en el uso de microsatélites y AFLP para el estudio poblacional de diversas especies de peces en el río Sinú, Colombia. Rev. colomb. biotecnol. 2004, 6, 72-78.
ABNT
LAMPREA, N.; LÓPEZ, L.; SANTACRUZ, D.; GUERRERO, J.; BURBANO, C. Modificaciones técnicas en el uso de microsatélites y AFLP para el estudio poblacional de diversas especies de peces en el río Sinú, Colombia. Revista Colombiana de Biotecnología, [S. l.], v. 6, n. 1, p. 72–78, 2004. Disponível em: https://revistas.unal.edu.co/index.php/biotecnologia/article/view/584. Acesso em: 11 feb. 2026.
Chicago
Lamprea, Natalia, Liliana López, Diana Santacruz, Jimena Guerrero, y Consuelo Burbano. 2004. «Modificaciones técnicas en el uso de microsatélites y AFLP para el estudio poblacional de diversas especies de peces en el río Sinú, Colombia». Revista Colombiana De Biotecnología 6 (1):72-78. https://revistas.unal.edu.co/index.php/biotecnologia/article/view/584.
Harvard
Lamprea, N., López, L., Santacruz, D., Guerrero, J. y Burbano, C. (2004) «Modificaciones técnicas en el uso de microsatélites y AFLP para el estudio poblacional de diversas especies de peces en el río Sinú, Colombia», Revista Colombiana de Biotecnología, 6(1), pp. 72–78. Disponible en: https://revistas.unal.edu.co/index.php/biotecnologia/article/view/584 (Accedido: 11 febrero 2026).
IEEE
[1]
N. Lamprea, L. López, D. Santacruz, J. Guerrero, y C. Burbano, «Modificaciones técnicas en el uso de microsatélites y AFLP para el estudio poblacional de diversas especies de peces en el río Sinú, Colombia», Rev. colomb. biotecnol., vol. 6, n.º 1, pp. 72–78, ene. 2004.
MLA
Lamprea, N., L. López, D. Santacruz, J. Guerrero, y C. Burbano. «Modificaciones técnicas en el uso de microsatélites y AFLP para el estudio poblacional de diversas especies de peces en el río Sinú, Colombia». Revista Colombiana de Biotecnología, vol. 6, n.º 1, enero de 2004, pp. 72-78, https://revistas.unal.edu.co/index.php/biotecnologia/article/view/584.
Turabian
Lamprea, Natalia, Liliana López, Diana Santacruz, Jimena Guerrero, y Consuelo Burbano. «Modificaciones técnicas en el uso de microsatélites y AFLP para el estudio poblacional de diversas especies de peces en el río Sinú, Colombia». Revista Colombiana de Biotecnología 6, no. 1 (enero 1, 2004): 72–78. Accedido febrero 11, 2026. https://revistas.unal.edu.co/index.php/biotecnologia/article/view/584.
Vancouver
1.
Lamprea N, López L, Santacruz D, Guerrero J, Burbano C. Modificaciones técnicas en el uso de microsatélites y AFLP para el estudio poblacional de diversas especies de peces en el río Sinú, Colombia. Rev. colomb. biotecnol. [Internet]. 1 de enero de 2004 [citado 11 de febrero de 2026];6(1):72-8. Disponible en: https://revistas.unal.edu.co/index.php/biotecnologia/article/view/584
Descargar cita
Visitas a la página del resumen del artículo
713
Descargas
Los datos de descargas todavía no están disponibles.
Licencia
Derechos de autor 2004 Revista Colombiana de Biotecnología

Esta obra está bajo una licencia internacional Creative Commons Atribución 4.0.
Esta es una revista de acceso abierto distribuida bajo los términos de la Licencia Creative Commons Atribución 4.0 Internacional (CC BY). Se permite el uso, distribución o reproducción en otros medios, siempre que se citen el autor(es) original y la revista, de conformidad con la práctica académica aceptada. El uso, distribución o reproducción está permitido desde que cumpla con estos términos.
Todo artículo sometido a la Revista debe estar acompañado de la carta de originalidad. DESCARGAR AQUI (español) (inglés).















