



Publicado
Caracterización bioquímica de hojas de clones de Theobroma cacao y su relación con los tricomas
Biochemical characterization of leaves in Theobroma cacao clones and its relation with trichomes
DOI:
https://doi.org/10.15446/rev.colomb.biote.v17n2.54265Palabras clave:
análisis de componentes principales, fenoles, prolina, proteínas, carbohidratos, principal components analysis, phenols, proline, proteins, carbohydrates (es)Título en ingles: Biochemical characterization of leaves in Theobroma cacao clones and its relation with trichomes
Theobroma cacao L. es un cultivo de gran importancia por sus posibles aplicaciones en la farmacia y la agricultura. Contiene metabolitos que resultan de gran interés por sus funciones biológicas naturales en la respuesta de la planta a los diferentes tipos de estrés, tanto bióticos como abióticos. La presencia, tipo y concentración de los metabolitos (primarios y secundarios) pueden variar de acuerdo al genotipo analizado y se pueden acumular en estructuras asociadas a la arquitectura de la hoja como son los tricomas. El objetivo de este trabajo fue caracterizar 26 clones del banco de germoplasma de Padrón, INIA-Miranda, Estado Miranda, Venezuela y estudiar la relación existente entre los indicadores bioquímicos y el número de tricomas. Se determinó el contenido de compuestos fenólicos, proteínas solubles, prolina libre, carbohidratos totales, ligninas, el número de tricomas y la relación existente entre ellos. Los resultados mostraron que en el espacio bidimensional que explica el 46,68% de la variabilidad total, se definieron cinco grupos homogéneos: diez clones se asociaron con el máximo gradiente de prolina, carbohidratos totales y contenido de ligninas, nueve con alto contenido de proteínas y fenoles y siete con mayor número de tricomas en hojas. Se comprobó que el número de tricomas no se relacionó con la concentración de compuestos fenólicos.Los clones de cacao del tipo forastero mostraron mayor concentración de fenoles totales y carbohidratos, en tanto que los trinitarios presentaron mayor concentración de proteínas y prolina.
Theobroma cacao L. is a crop of major importance for their potential applications in pharmacy and agriculture. It contains metabolites that are of great interest for its natural biological functions in plant response to different types of stress, both biotic and abiotic. The presence, type and concentration of both primary and secondary metabolites may vary depending on the genotype analyzed and these compounds can accumulate in the architecture associated structures such as leaf trichomes. The aim of this study was to characterize 26 clones of the germplasm bank of Padrón, INIA-Miranda, Miranda State, Venezuela and study the relationship between biochemical indicators assessed and the number of trichomes. The content of phenolic compounds, soluble protein, free proline, total carbohydrates, lignins, the number of trichomes and the relationship between all traits was determined. The results showed that in the two dimensional space explained 46.68% of the variability, five homogeneous groups were defined: 10 clones were associated with the maximum gradient of proline, total carbohydrate and lignin content. Nine clones were associated with high content of phenols and a group of seven clones with the highest number of trichomes on leaves. It was found that the number of trichomes was not related to the concentration of phenolic compounds. The clones of Forastero cocoa type showed higher concentration of total phenols and carbohydrates, meanwhile the Trinitario type presented higher concentration of proteins and prolina.
Key words: principal components analysis; phenols; proline; proteins; carbohydrates.
DOI: https://doi.org/10.15446/rev.colomb.biote.v17n2.54265
ARTÍCULO DE INVESTIGACIÓN
Caracterización bioquímica de hojas de clones de Theobroma cacao y su relación con los tricomas
Biochemical characterization of leaves in Theobroma cacao clones and its relation with trichomes
Janet Quiñones-Galvez*1, Daynet Sosa**1,2, JR. Demey***, Silvia Alemán****, Maryla Sosa****; Dercy Parra*****, Olga Móvil*****, Reinaldo Trujillo*, Yanelis Capdesuñer*, Yemeys Quirós*, Martha Hernández*3, Diógenes Infante******3
* MSc. Janet Quiñones-Galvez, Dra. Martha Hernández, Dr. Reinaldo Trujillo, MSc. Yanelis Capdesuñer, Ing. Yemeys Quirós, Centro de Bioplantas, Universidad de Ciego de Ávila, Carretera a Morón km 9 ½. Ciego de Ávila, Cuba. Email: jquinones@bioplantas.cu, janet.quinonesgalvez@gmail.com.
** Dra. Daynet Sosa, Fundación Instituto de Estudios Avanzados IDEA, Carretera Nacional Hoyo de la Puerta. Valle Sartenejas. Baruta. Caracas, Estado Miranda, Venezuela. Universidad Estatal de Milagro, UNEMI, Milagro; Centro de Investigaciones Biotecnológicas del Ecuador, CIBE, Escuela Superior Politécnica del Litoral, ESPOL, Guayaquil; Guayas, Ecuador (Dirección actual).
*** Dr. JR. Demey, Fundación Instituto de Estudios Avanzados IDEA, Carretera Nacional Hoyo de la Puerta. Valle Sartenejas. Baruta. Caracas, Estado Miranda, Venezuela. Escuela Superior Politécnica del Litoral, ESPOL, Facultad de Ciencias Naturales y Matemáticas, Guayaquil, Ecuador (Dirección actual).
**** MSc. Silvia Alemán, MSc. Maryla Sosa. Universidad de Matanzas, Matanzas, Cuba.
***** MSc. Dercy Parra, MSc. Olga Móvil. Estación Experimental de Padrón, INIA-Miranda, Caucagua, Estado Miranda, Venezuela.
****** Dr. Diógenes Infante, Fundación Instituto de Estudios Avanzados IDEA, Carretera Nacional Hoyo de la Puerta. Valle Sartenejas. Baruta. Caracas, Estado Miranda, Venezuela.
1 Ambas autoras tuvieron igual porcentaje de participación, por lo que se consideran autores principales.
2 Codirectora del proyecto de investigación
3 Directores del proyecto de investigación
Recibido: febrero 18 de 2015 Aprobado: septiembre 18 de 2015
Resumen
Theobroma cacao L. es un cultivo de gran importancia por sus posibles aplicaciones en la farmacia y la agricultura. Contiene metabolitos que resultan de gran interés por sus funciones biológicas naturales en la respuesta de la planta a los diferentes tipos de estrés, tanto bióticos como abióticos. La presencia, tipo y concentración de los metabolitos (primarios y secundarios) pueden variar de acuerdo al genotipo analizado y se pueden acumular en estructuras asociadas a la arquitectura de la hoja como son los tricomas. El objetivo de este trabajo fue caracterizar 26 clones del banco de germoplasma de Padrón, INIA-Miranda, Estado Miranda, Venezuela y estudiar la relación existente entre los indicadores bioquímicos y el número de tricomas. Se determinó el contenido de compuestos fenólicos, proteínas solubles, prolina libre, carbohidratos totales, ligninas, el número de tricomas y la relación existente entre ellos. Los resultados mostraron que en el espacio bidimensional que explica el 46,68% de la variabilidad total, se definieron cinco grupos homogéneos: diez clones se asociaron con el máximo gradiente de prolina, carbohidratos totales y contenido de ligninas, nueve con alto contenido de proteínas y fenoles y siete con mayor número de tricomas en hojas. Se comprobó que el número de tricomas no se relacionó con la concentración de compuestos fenólicos. Los clones de cacao del tipo forastero mostraron mayor concentración de fenoles totales y carbohidratos, en tanto que los trinitarios presentaron mayor concentración de proteínas y prolina.
Key words: análisis de componentes principales; fenoles; prolina; proteínas; carbohidratos.
Abstract
Theobroma cacao L. is a crop of major importance for their potential applications in pharmacy and agriculture. It contains metabolites that are of great interest for its natural biological functions in plant response to different types of stress, both biotic and abiotic. The presence, type and concentration of both primary and secondary metabolites may vary depending on the genotype analyzed and these compounds can accumulate in the architecture associated structures such as leaf trichomes. The aim of this study was to characterize 26 clones of the germplasm bank of Padrón, INIA-Miranda, Miranda State, Venezuela and study the relationship between biochemical indicators assessed and the number of trichomes. The content of phenolic compounds, soluble protein, free proline, total carbohydrates, lignins, the number of trichomes and the relationship between all traits was determined. The results showed that in the two dimensional space explained 46.68% of the variability, five homogeneous groups were defined: 10 clones were associated with the maximum gradient of proline, total carbohydrate and lignin content. Nine clones were associated with high content of phenols and a group of seven clones with the highest number of trichomes on leaves. It was found that the number of trichomes was not related to the concentration of phenolic compounds. The clones of Forastero cocoa type showed higher concentration of total phenols and carbohydrates, meanwhile the Trinitario type presented higher concentration of proteins and prolina.
Key words: principal components analysis; phenols; proline; proteins; carbohydrates.
Introducción
Los metabolitos son los responsables de muchos fenómenos y funciones indispensables para el crecimiento y desarrollo de plantas, así como de su interacción con el medio ambiente que las rodea. Dentro de ellos, se encuentran compuestos de gran importancia para el crecimiento y supervivencia de las plantas como fenoles, proteínas, prolina y carbohidratos (Petti y Scully, 2009; Yooyongwech et al., 2012; Mulekar y Huq, 2014; Van den Ende, 2014).
Los compuestos fenólicos son metabolitos secundarios de naturaleza compleja en las plantas, se relacionan con los procesos de maduración, mecanismos de defensa, respuesta a estrés biótico y abiótico, y a su vez tienen gran influencia en las propiedades organolépticas de los productos alimenticios derivados de plantas (Kubo y Matsumoto, 1984; Cimato et al., 1990; Niemenak et al., 2006). Mientras que las proteínas, la prolina y los carbohidratos son metabolitos primarios que pueden estar involucrados, al igual que los fenoles, en mecanismos de resistencia a estrés biótico y abiótico, como es el caso de las proteínas relacionadas con la patogénesis (PR) (Martinez-Esteso et al., 2009), la prolina que se relaciona con la respuesta a sequía en algunas plantas (Wagner et al., 2003), los carbohidratos de la superficie de la hoja tienen un papel importante en la colonización temprana y crecimiento de hongos, así como la sacarosa favorece la formación de biopelículas de bacterias en medio de cultivo (Lindow y Brandl, 2003; Tomihama et al., 2006).
El Theobroma cacao L. (T. cacao L.) se cultiva a lo largo de las zonas tropicales del planeta (Elwers et al., 2009) aunque es una planta nativa de la región tropical de las Américas (Rusconi y Conti, 2010). Se utiliza en la medicina popular como antiséptico, diurético y parasiticida (Edwards et al., 2005) y tales propiedades se asocian a los variados metabolitos que contiene. Los mismos pueden encontrarse en diferentes órganos de la planta (hojas, tallos, raíces, flores, semillas, tricomas) de acuerdo a su ruta biosintética y su función biológica (Ávalos y Pérez-Urria, 2009). Actualmente existen estudios encaminados al análisis de la calidad y sabor del chocolate por la caracterización de los cambios en la composición de proteínas y fenoles durante la maduración de los frutos de cacao, la fermentación de las semillas y pasos claves en la producción industrial del chocolate (Pirovani et al., 2008; Guehi et al., 2010; Afoakwa et al., 2012a; Afoakwa et al., 2012b).
Las plantas con mayores concentraciones de proteínas revisten gran interés debido a su importancia para el desarrollo de las plantas, así como para el beneficio humano. Mientras que los fenoles se conocen también por su beneficio a la salud humana y se les han encontrado diversas actividades, entre las que se encuentran: moduladores de la absorción de grasas, anti-inflamatorios, en la reducción de enfermedades cardiovasculares al reducir la síntesis de lípidos y como antioxidantes (Guerrero et al., 2013; Gu et al., 2014; Sarria et al., 2014). Por otra parte se les conoce por su actividad antimicrobiana contra patógenos que afectan a las plantas y al hombre (Godard et al., 2009; Li et al., 2009; Vivot et al., 2009; Mishra et al., 2010; Pandey y Mishra 2010).
La superficie de hojas de la mayoría de las plantas está cubierta por células modificadas de la epidermis que se denominan tricomas (Peter y Shanower, 1998). Los compuestos que se encuentran en los tricomas de la superficie foliar, pueden estar involucrados fundamentalmente en la defensa a diferentes condiciones de iluminación y ataque de patógenos e insectos (Wagner, 1991; Peter y Shanower, 1998; Wagner et al., 2004; Ishida et al., 2008). En plantas de cacao, se encontró que los tricomas grandulares pueden proveer el punto de entrada de especies de Tricoderma en el tallo, contribuyendo a una colonización sistémica del tejido y al establecimiento de una asociación endofítica (Bailey et al., 2009).
Por todo lo anterior, y hasta donde tenemos conocimiento, falta información en cuanto a las similitudes y diferencias bioquímicas, así como la relación que existe entre los tricomas y la composición química de los clones de T. cacao L. que se encuentran en el banco de germoplasma de Padrón, INIA-Miranda, Venezuela. En este sentido, el objetivo de esta investigación fue estudiar la relación existente entre los indicadores bioquímicos (contenido de compuestos fenólicos, proteínas solubles, prolina libre, carbohidratos totales, ligninas) y el número de tricomas, para caracterizar el comportamiento en 26 clones de dicho banco.
Materiales y métodos
Material vegetal
Las muestras se colectaron en el Banco de Germoplasma de la Estación Experimental de Padrón, INIA-Miranda, Estado Miranda, Venezuela (tabla 1).

Las muestras de T. cacao se colectaron en el Banco de Germoplasma de la Estación Experimental de Padrón, INIA-Miranda, Estado Miranda, Venezuela. Ubicado a 41 m s.n.m., a 10º13'14.1"N y 66º17'57.5"W. Las plantas muestreadas tenían 12 años de plantadas en bosque húmedo tropical, en un suelo franco-arcilloso, con pH 5,3, topografía plana y sin irrigación. La densidad de plantación fue de 1600 plantas.ha-1 y la fertilización con 100 g NPK (12-12-17) + 50 g urea. La humedad osciló entre 72-89%, con un promedio de lluvias de 2441 mm.
Para los análisis bioquímicos y el conteo de tricomas, se tomaron las cuatro hojas verdes más jóvenes, de ramas plagiotrópicas de edad media de acuerdo al número de ramas, de tres plantas de cada clon evaluado de T. cacao L., inmediatamente se colocaron en nitrógeno líquido y se almacenaron a -80 °C hasta la realización de las extracciones.
Compuestos fenólicos
La extracción y determinación de fenoles se desarrolló de acuerdo al método de Friend, (1992) con modificaciones. Para ello, las hojas (limbos foliares, sin pecíolo ni nervio central) se maceraron en nitrógeno líquido hasta polvo fino (0,1 g por triplicado de la mezcla de hojas de cada material vegetal) y se extrajeron tres veces con metanol hasta volumen final 1mL (fenoles solubles). Los residuos se secaron y se hidrolizaron con 0,25 mL de NaOH 2 mol.L-1, 16 h a 70 ºC. Posteriormente se adicionó 0,25 mL HCl 2 mol.L-1. La mezcla se centrifugó a 15 000 g por 5 min y se colectó el sobrenadante (fenoles ligados a la pared). El residuo se lavó con 7 mL de agua destilada, se ajustó a pH 7,5 (con NaOH/HCl) y se enrazó a 10 mL con agua destilada (ligninas).
Los fenoles solubles y ligados a la pared se determinaron con el reactivo Folin-Ciocalteu. Una alícuota de cada extracto se diluyó a 1 mL con agua destilada y se adicionó 0,1 mL de reactivo Folin-Ciocalteu. Después de 5 min se adicionó 0,6 mL de Na2CO3 saturado en NaOH 1 mol.L-1. Se incubó 60 min en la oscuridad. La medición de la coloración desarrollada se realizó en espectrofotómetro a 725 nm. La concentración de fenoles se expresó en mg. g-1 de masa fresca (MF), equivalente a una curva patrón de ácido clorogénico. Los fenoles totales se calcularon con la sumatoria de los fenoles solubles y los ligados a la pared.
Para la cuantificación de ligninas se añadió 4 mL de NaOH 0,1 mol.L-1 (pH 12) en 1 mL de cada extracto y 4 mL de buffer fosfato (pH 7) en otro mL. La absorbancia se midió 350 nm, extracto a pH 12 contra el de pH 7. Los resultados se expresaron como unidades de absorbancia (UA) por 0,1 g MF.
Proteínas solubles
Para la extracción de las proteínas solubles, se partió de hojas maceradas de igual forma que para los fenoles. Se añadió 1 mL de buffer de extracción (Urea: 6 mmol.L-1, EDTA: 10 mmol.L-1, NaH2PO4: 0,1 mol.L-1, β-mercaptoetanol: 14 mmol.L-1, Tween-20 (0,05 %), Tris HCl: 10 mmol.L-1 pH= 8) a 0,1 g de material vegetal (triplicado de la mezcla de hojas de cada material vegetal). La mezcla se homogenizó y se centrifugó en frío a 4ºC por 10 min. El sobrenadante se colectó para la cuantificación. La concentración de PST se determinó de acuerdo a Bradford, (1976) y la medición de la coloración desarrollada se realizó en espectrofotómetro a 595 nm. La concentración se expresó en mg.g-1 MF, equivalente a una curva patrón de albúmina de suero bovino.
Carbohidratos totales y prolina libre
La extracción de carbohidratos y prolina libre se realizó a hojas maceradas con el procedimiento de los fenoles. Se agregó 10 mL de metanol: agua 80% (v:v) a 1 g de material vegetal (triplicado de la mezcla de hojas de cada material vegetal). La mezcla se agitó con vórtex y se centrifugó a temperatura ambiente a 4000 g por 8 min. La extracción se repitió 2 veces. Los sobrenadantes se colectaron, unificaron y filtraron con papel de filtro. La fase etanólica se eliminó a vacío en roto-evaporador. La fase acuosa se colectó para las cuantificaciones.
La concentración de carbohidratos se determinó de acuerdo al método de Dubois et al., (1956). A 1 mL de extracto, se adicionó 0,5 mL de mezcla fenol: agua 5% (v:v) y 2,5 mL de H2SO4 concentrado cuidadosamente. Se incubó por 30 min a temperatura ambiente. La medición de la coloración desarrollada se realizó en espectrofotómetro a 492 nm. La concentración de carbohidratos totales se expresó en µg.g-1 MF, equivalente a una curva patrón de glucosa.
La concentración de prolina libre se determinó de acuerdo a Bates et al., (1973). A 0,5 mL del extracto vegetal se adicionó 0,5 mL de ninhidrina y 0,5 mL de ácido acético. Posteriormente se agitó y colocó en baño de maría a 100°C por 1h. Se agregó 1 mL de tolueno y se agitó fuertemente. La absorbancia se determinó a la fase orgánica a 520 nm. La concentración se expresó en µg.g-1 MF, equivalente a una curva patrón de L-prolina.
Conteo de tricomas
El número y tipo de tricomas por campo visual se contó directamente en microscopio óptico, partiendo de la lámina epidérmica de la cara abaxial de las hojas de los 26 clones (seis réplicas del total de hojas de cada material vegetal). Las evaluaciones se realizaron en microscopio LEICA DM 3000 con lente 20x.
Análisis estadístico
El procesamiento estadístico de los resultados se realizó con el utilitario Statistical Package for Social Sciences (SPSS) (versión 20 para Windows, SPSS Inc.). Se realizaron pruebas paramétricas (ANOVA de un factor y bifactorial, Tuckey). Para estudiar la relación entre el número de tricomas y la composición bioquímica de las hojas, se utilizó el Análisis de Componentes Principales (ACP). Este análisis se realizó con InfoStat (versión 2011, Grupo InfoStat, FCA, Universidad Nacional de Córdoba, Argentina). Los detalles del tratamiento estadístico aparecen en cada figura o tabla de resultados y discusión.
Resultados y discusión
La concentración de fenoles solubles, fenoles ligados a la pared, lignina, proteínas solubles y prolina libre en 26 clones de T. cacao se muestra en la tabla 2.
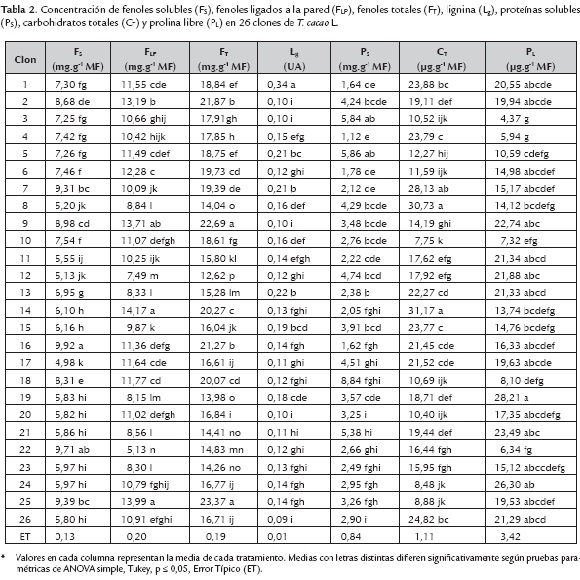
Para los fenoles solubles, los clones 16 y 22 mostraron, sin diferencias significativas, los mayores valores con 9,92 y 9,71 mg.g-1 MF respectivamente y el clon 17 la menor, con 4,98 mg.g-1 MF. Mientras que en los clones 9, 14 y 25 se encontraron la mayor concentración de fenoles ligados a la pared, con 13,71; 14,17 y 13,99 mg.g-1 MF respectivamente y sin diferencias significativas, el menor valor se encontró en el clon 22. Al analizar los fenoles totales, los clones 9 y 25 fueron los superiores, sin diferencias significativas entre ellos y con una concentración de 22,69 y 23,37 mg.g-1 MF, le siguen los clones 2 y 16 con 21,87 y 21,27 mg.g-1 MF y posteriormente el 6, 14 y 18, con 19,73; 20,27 y 20,07 respectivamente.
Se puede apreciar además, que para las ligninas fue el clon 1, con 0,35 UA, el de mayor contenido de ligninas y los menores valores estuvieron en los clones: 2, 3, 9, 20 y 26, con 0,10; 0,10; 0,10; 0,10 y 0,09 UA respectivamente y sin diferencias significativas. Estos resultados indican una mayor concentración de compuestos fenólicos ligados a la pared que los solubles y no coinciden los clones que tienen mayor concentración de ambos tipos de fenoles. Los compuestos fenólicos son parte importante de la defensa activa de plantas. Estos compuestos se pueden sintetizar y movilizar en condiciones de estrés y en ocasiones se esterifican para formar parte de la pared celular y de lignina (Fry, 1987; Nicholson y Hammerschmidt, 1992; Cabrera-Soto et al., 2009). En este caso, todas las plantas estaban expuestas a condiciones similares de cultivo y sin síntomas de enfermedades, por lo que no se puede afirmar que las diferencias en la composición de fenoles estén asociadas a respuesta a un determinado tipo de estrés.
Se puede apreciar que para el caso de las proteínas solubles los clones: 3, 5 y 18 mostraron las mayores concentraciones, con 5,84; 5,86 y 8,84 mg.g-1 MF, los menores se encontraron para el clon 4, con 1,12 mg.g‑1 MF. En los carbohidratos totales las mayores concentraciones se encontraron en los clones: 7, 8 y 14, con 28,13; 30,73 y 31,17 µg.g-1 MF respectivamente, mientras que la menor concentración fue para el clon 10, con 7,75 µg.g-1 MF. Al analizar la prolina libre, los clones 19 y 24 fueron superiores, con 28,22 y 26, 29 µg.g-1 MF, mientras que los clones: 3, 4 y 22 mostraron menores concentraciones con 4, 37; 5,94 y 6,34 µg.g-1 MF.
En cuanto al número y tipo de tricomas, en la figura 1 se puede observar que se encontraron tricomas glandulares (señalados en figura 1 a, g, h, i), no glandulares estrellados (señalados en figura 1 a, b, c, i) y no glandulares simples (señalados en figura 1 d, e, f).
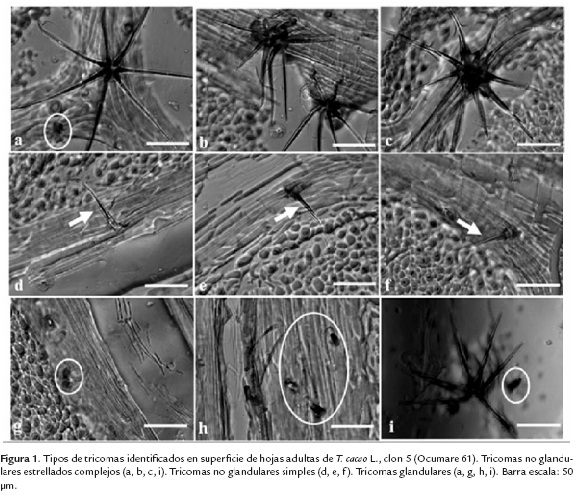
No hubo diferencias en el tipo de tricomas para los 26 los clones evaluados, se observó un comportamiento similar en todos los casos, con tricomas glandulares, no glandulares estrellados y no glandulares simples, aunque el tipo más frecuente fue el no glandular estrellado.
Como se puede observar en la figura 1 (a-i), los tricomas de cacao mostraron diversidad en cuanto a número y tipo, así como en las ramificaciones de los no glandulares estrellados. Se encontró además, la acumulación de compuestos coloreados dentro de las mismas, los cuales pueden ser fenólicos u otros metabolitos de interés como terpenos, entre otros (Tattini et al., 2000, McDowell et al., 2011). No obstante, para corroborar el tipo de compuestos presente es necesario recurrir a otros tipos de microscopía.
El número de tricomas varió en dependencia del clon, aunque la mayoría de los clones no mostraron diferencias significativas. La mayor cantidad de tricomas se encontró en los clones 5, 23 y 26, con 45, 40 y 40 tricomas. mm-2, sin diferencias significativas, y la menor en el clon 10, con 15 tricomas mm-2. En hojas de papa y tomate, los tricomas contienen fenoles y enzimas oxidadoras de fenoles (polifenol oxidasas) que pueden matar insectos, dentro de los que se encuentran los áfidos (Peter y Shanower, 1998). Es por ello que resulta de gran interés agrupar los clones por sus compuestos mayoritarios, así como evaluar si existe relación entre el número de tricomas y los compuestos químicos de las hojas de T. cacao.
En la figura 2 se muestra el análisis de la relación entre los 26 clones estudiados de T. cacao, su contenido de prolina libre, proteínas solubles, carbohidratos totales, ligninas, fenoles solubles, fenoles ligados a la pared y el número de tricomas.
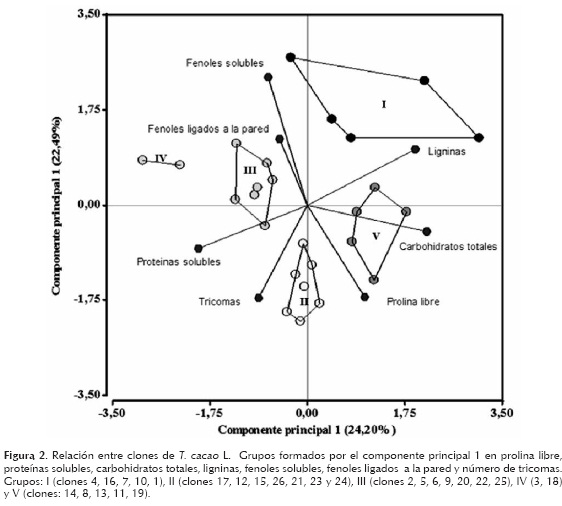
Los resultados muestran que en el espacio bidimensional, que explica el 46,68% de la variabilidad total, se definieron cinco grupos homogéneos. El grupo 1 (I), formado por los clones: 4, 16, 7, 10, 1, asociados a la máxima concentración de ligninas. El grupo 2 (II) formado por los clones 17, 12, 15, 26, 21, 23 y 24, asociado a plantas con hojas con el mayor número de tricomas. Los grupos 3 (III) y 4 (IV) formados por los clones: 2, 5, 6, 9, 20, 22, 25; y 3, 18 respectivamente, asociados a altos contenidos de proteínas solubles, fenoles solubles y ligados a la pared y el grupo 5 (V), formado por los clones: 14, 8, 13, 11, 19 respectivamente, asociados al gradiente de máximo contenido de prolina libre y carbohidratos totales.
Al comparar los tipos de cacao (forastero y trinitario), en cuanto a la concentración de fenoles totales, lignina, proteínas solubles y prolina libre (tabla 3) se encontró que los clones trinitarios tienen mayor contenido de proteínas solubles y prolina libre que los forasteros. Mientras que estos últimos superan a los trinitarios en fenoles totales (fenoles solubles + fenoles ligados a la pared) y carbohidratos.
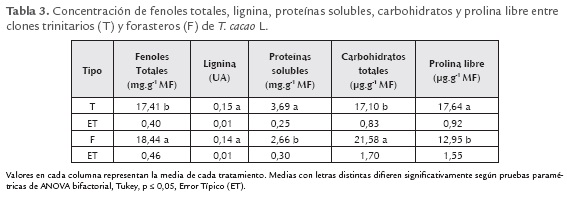
Es interesante señalar que: las sustancias producidas por las partes aéreas de las plantas pueden ser carbohidratos, aminoácidos orgánicos, fenoles y otros compuestos, dentro de los fenoles están los taninos, que generalmente son toxinas por su capacidad de unirse a proteínas y actúan como repelentes alimenticios de muchos animales que pueden ser dañinos a las plantas, sin embargo pueden tener efecto positivo en la salud humana (Ávalos y Pérez-Urria, 2009). Para el caso de los fenoles totales (tabla 3), su concentración superior en clones forasteros puede estar relacionado con que este tipo de cacao es más resistente y de menor calidad para la industria por su alto contenido de fenoles, dentro de los que se encuentran los taninos, con una mayor concentración en estos clones (Figueira et al., 1997).
Las diferencias cuantitativas encontradas en el contenido de fenoles (tabla 2) entre los clones, se asocian mayormente al genotipo, debido a que las plantas tenían igual edad y estaban cultivadas en similares condiciones, lo que coincide con lo planteado por Zapata et al. (2000) que demostró, en tabaco, que los fenoles dependen de la edad, el estado de desarrollo y el genotipo. Niemenak et al., (2006) encontraron que los fenoles totales, catequina, epicatequina y antocianina en semillas frescas y fermentadas de T. cacao L., fueron dependientes del genotipo pero no observaron diferencias cualitativas en el contenido de polifenoles en relación con su origen genético, las diferencias se relacionaron más con las condiciones de crecimiento. Sin embargo, en el presente trabajo se encontró diferencias en la concentración de fenoles, carbohidratos, proteínas y prolina asociadas al genotipo y también al origen genético, ya que en los forasteros la concentración de fenoles totales y carbohidratos totales fue superior a los trinitarios (tabla 3), en tanto que los trinitarios fueron mayoritarios en proteínas solubles y la prolina libre. Además, todos los indicadores evaluados variaron entre los diferentes clones (tabla 3).
Coincidiendo con Chen et al., (2005) plantean que las proteínas solubles, la ribulosa-1,5-bisfosfato carboxilasa/oxigenasa (Rubisco) y su contenido activasa, así como la actividad de la sacarosa fosfato sintasa, decrecen significativamente, mientras que el contenido de azúcares solubles, fosfato inorgánico, clorofilas y el complejo II de la fotosíntesis incrementaron en hojas de arroz aclimatizadas con aire enriquecido con CO2. En la actualidad se conoce que las proteínas se encuentran en baja concentración en los tejidos vegetales en comparación con animales y bacterias, además en plantas maduras son más difíciles de extraer por el espesor de la pared celular (Islam et al., 2004). Por otra parte, los compuestos fenólicos combinan con las proteínas por condensaciones covalentes (Carpentier et al., 2005) y las proteínas como la Rubisco son muy abundantes, lo que puede dificultar la detección de las que están en menor abundancia (Neilson et al., 2010; Balbuena et al., 2011).
Matt et al., (2002) transformaron plantas de tabaco con una construcción antisentido de Rubisco para estudiar las consecuencias de una lesión en el metabolismo del carbono fotosintético en el metabolismo del nitrógeno y el metabolismo secundario. Ellos demostraron que la inhibición de la fotosíntesis y el decrecimiento del contenido de azúcares conllevan a la inhibición del metabolismo de nitrógeno y a cambios dramáticos en la composición de los metabolitos secundarios de las hojas, como ácido clorogénico y nicotina, que son ricos en carbono y nitrógeno en su estructura, respectivamente. Demostraron además que los cambios en la razón aminoácido: azúcares se relacionaron con cambios en la razón nicotina: ácido clorogénico. Estos resultados se corresponden con los presentados en la tabla 3, donde en el grupo que presentó mayor concentración de fenoles (clones forasteros), se encontró también mayor concentración de carbohidratos.
Por otra parte, las proteínas, carbohidratos y prolina juegan papeles importantes en la respuesta al estrés, donde se ha demostrado que en hojas infectadas por C. perniciosa decrecen las proteínas solubles (Frias et al., 1991). También se ha encontrado que en hojas de plantas de T. grandiflorum infectadas por hongo, se afecta el contenido de clorofilas y por lo tanto se afecta también la fotosíntesis y la síntesis de carbohidratos, así como se producen variaciones en las concentraciones de fenoles (Oliveira et al., 1997).
Como se puede observar, los grupos que se forman con el ACP no se relacionan con el tipo de cacao, pues los clones forasteros y trinitarios quedan mezclados dentro de los diferentes grupos (figura 2). En la tabla 3, el grupo de los clones forasteros mostraron mayor concentración de carbohidratos y fenoles, los trinitarios en prolina y proteínas, y no existieron diferencias entre los dos grupos para ligninas. Sin embargo, en el ACP los clones se agruparon de otra manera: cinco clones, se asociaron por su alto contenido de ligninas (tres forasteros y dos trinitarios), otros cinco por la alta concentración de prolina y carbohidratos totales (dos forasteros y tres trinitarios), nueve por su alto contenido de proteínas y fenoles (todos trinitarios) y siete con mayor número de tricomas (un forastero y seis trinitarios). Contrario a lo esperado, en el análisis de ACP, el mayor número de estructuras acumuladoras de metabolitos (tricomas) se relacionó más con la mayor concentración de proteínas solubles y la prolina libre que con el resto de los parámetros evaluados.
En el caso de los tricomas, se conoce que la combinación de tricomas glandulares y no glandulares confiere resistencia a diferentes plagas, debido a que los glandulares contienen o secretan mezclas de compuestos químicos y los no glandulares sirven de barrera mecánica sobre todo para los insectos (Peter y Shanower, 1998). Los clones con mayor número de fenoles no resultaron asociados a los mayores contenidos de tricomas. Lo cual no descarta el hecho de que existan tipos de fenoles que se encuentren en los tricomas, o secretados por ellos y que sean de importancia por su actividad biológica, lo que los haría interesantes para la farmacia, control de patógenos y obtención de pigmentos o aromatizantes. No obstante, para seleccionar clones por la producción de compuestos en los tricomas habría que evaluar no solo cuáles son los de mayor número, sino el perfil de metabolitos presentes en los tricomas de cada clon y el valor de los mismos por su actividad.
En plantas del género Solanum existen seis tipos de tricomas y en todos los de tipo 1, 4 y 6 se encontraron genes para la biosíntesis de metabolitos como flavonoides, azúcares acetilados y terpenoides (McDowell et al., 2011). Hasta ahora en cacao no se ha descrito que tipo de compuestos pueden estar en los tricomas, pero si se conoce que hay tricomas glandulares y no glandulares (Bailey et al., 2009) y que además hay un alto contenido de fenoles, por lo que también podría existir síntesis y secreción de compuestos similares a los descritos para Solanum.
Finalmente, los clones analizados en la presente investigación no estaban visiblemente infectados por ningún patógeno, no obstante, dentro de los 26 clones existen algunos con mayor resistencia a patógenos que otros. Por lo que las concentraciones de los compuestos analizados pueden estar apuntando a mayor cantidad de ellos en clones resistentes que en los susceptibles a enfermedades fungosas. Por ejemplo, en investigaciones previas realizadas en el banco de germoplasma del que se seleccionó el material evaluado, el clon 3 (alto contenido de proteínas y fenoles) es resistente a P. palmivora y a C. frimbriata, el clon 8 (alto contenido de carbohidratos y prolina) es moderadamente resistente a P. palmivora, el clon 4 (alto contenido de ligninas) es resistente a C. frimbriata y tiene baja incidencia de M. roreri, el clon 24 (mayor número de tricomas) es resistente a P. palmivora (Reyes et al., 1973; González et al., 2009; González et al., 2011; Morillo et al., 2012). Además, cuando el 19 (alto contenido de carbohidratos y prolina) fue utilizado como madre en dos cruces (19 x 4 y 19 x 22), la descendencia de ambos mostraron baja incidencia de P. palmivora en frutos cosechados y baja incidencia de M. perniciosa como escobas vegetativas y en cojines florales. Estos cruces presentaron buenos índices de productividad, valores de índice de almendra > 1,3 y de índice de mazorca < 22 (González et al., 2011). Lo que corrobora que cada grupo obtenido ofrece la posibilidad del manejo de sus clones en dependencia de su composición química y el futuro uso.
Conclusiones
Este estudio demostró que existen cinco grupos, de acuerdo a la relación existente entre la concentración de compuestos bioquímicos y los tricomas de hojas de 26 clones de T. cacao. Cinco clones, se asociaron por su alto contenido de ligninas, otros cinco por la alta concentración de prolina y carbohidratos totales, nueve por su alto contenido de proteínas y fenoles y siete con mayor número de tricomas, de acuerdo al ACP que explica el 46,68% de la variabilidad total. Además, el grupo de los clones forasteros mostró mayor concentración de fenoles totales y carbohidratos, en tanto que los trinitarios presentaron mayor concentración de proteínas y prolina. Los resultados obtenidos pueden servir de base para la selección de clones, teniendo en cuenta el contenido de los diferentes parámetros evaluados, en dependencia del interés de las futuras investigaciones.
Agradecimientos
Los autores agradecemos el financiamiento otorgado al proyecto "Metabolitos Secundarios en Theobroma cacao L.", en el marco de la Instrumentación de la Alternativa Bolivariana para los Pueblos de Nuestra América, a partir de la Modificación y Ampliación del Convenio Integral de Cooperación Cuba – Venezuela "VII Comisión Mixta", que permitió realizar la investigación y procesamiento estadístico de los datos.
Referencias
Afoakwa, E. O., Jennifer, Q., Agnes, S. B., Jemmy, S. T., y Firibu, K. S. (2012a). Influence of pulp-preconditioning and fermentation on fermentative quality and appearance of Ghanaian cocoa (Theobroma cacao) beans. International Food Research Journal, 19(1), 127-133.
Afoakwa, E. O., Quao, J., Takrama, F. S., Budu, A. S., y Saalia, F. K. (2012b). Changes in total polyphenols, o-diphenols and anthocyanin concentrations during fermentation of pulp pre-conditioned cocoa (Theobroma cacao) beans. International Food Research Journal, 19 (3), 1071-1077.
Ávalos, A., y Pérez-Urria, E. (2009). Metabolismo secundario de plantas. Reduca (Biología). Serie Fisiología Vegetal,2 (3), 119-145.
Bailey, B.A, Strem, M.D., y Wood, D. (2009). Trichoderma species form endophytic associations within Theobroma cacao trichomes. Mycological Research,113, 1365-1376.
Balbuena, T.S., Dias, L.L.C., Martins, M.L.B., Chiquieri, T.B., Santa-Catarina, C., Floh, E.I.S., y Silveira, V. (2011). Challenges in proteome analyses of tropical plants. Brazilian Journal of Plant Physiology,23(2), 91-104.
Bates, L.S., Waldron, R.P., y Teare, L.D. (1973). Rapid determination of free proline for water-stress studies. Plant Soil,39, 205–207.
Bradford, M.M. (1976). A rapid and sensitive method for quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Analytical Biochemistry,72, 248-254.
Cabrera-Soto, M.L., Salinas-Moreno, Y., Velázquez-Cardelas, G.A., y Espinosa-Trujillo, E. (2009). Content of soluble and insoluble phenols in the structures of corn grain and their relationship with physical properties. Agrociencia,43, 827-839.
Carpentier, S.C., Witters, E., Laukens, K., Deckers, P., Swennen, R., y Panis, B. (2005). Preparation of protein extracts from recalcitrant plant tissues: An evalution of different methods for two-dimensional gel electrophoresis analysis. Proteomics,5, 2497-2507.
Chen, G.Y., Yong, Z.H., Liao, Y., Zhang, D.Y., Chen, Y., Zhang, H.B., Chen, J., Zhu, J.G., y Xu, D.Q. (2005). Photosynthetic Acclimation in Rice Leaves to Free-air CO2 Enrichment Related to Both Ribulose-1,5-bisphosphate Carboxylation Limitation and Ribulose-1,5-bisphosphate Regeneration Limitation. Plant Cell Physiology, 46 (7), 1036-1045.
Cimato, A., Mattei, A., y Osti, M. (1990). Variation of polyphenol composition with harvesting period. Acta Horticulturae,286, 453-456.
Dubois, M., Gilles, K.A., Hamiltion, JK., Rebers, PA., y Smith, F. (1956). Colorimetric method for determination of sugars and related substances. Analytical Chemistry,28, 350-356.
Edwards, H.G.M., Villar, J.S.E., de Oliveira, L.F.C., y Le Hyaric, M. (2005). Analytical Raman spectroscopic study of cacao seeds and their chemical extracts. Analytica ChimicaActa,538, 175-180.
Elwers, S., Zambrano, A., Rohsius, Ch., y Lieberei, R. (2009). Differences between the content of phenolic compounds in criollo, forastero and trinitario cocoa seed (Theobroma cacao L.). European Food Research and Technology, 2009, 937-948.
Figueira, A., Lambert S., Carpenter, D., Pires, J., Cascardo, J., y Romanczyk, L. (1997). The similarity of cocoa flavour of fermented seeds from fingerprinted genotypes of Theobroma cacao L. from Brazil and Malaysia. Tropical Agriculture,74(2), 132-139.
Friend, J. (1992). Lignin and associated phenolic acids in cell walls. In: Gurr, S.J., McPherson, M.J., Bowles, D.J. (eds.), Molecular plant pathology: a practical approach. Oxford: IRL Press, 2, pp. 51- 59.
Fry, S.C. (1987). Intercellular feruloylation of pectic polysaccharides. Planta, 171, 205-211.
Godard, S., Slacanin, I., Viret, O., y Gindro, K. (2009). Induction of defense mechanisms in grapevine leaves by emodin- and anthraquinone-rich plant extracts and their conferred resistance to downy mildew. Plant Physiology and Biochemistry,47, 827-837.
González, R.V., Girón, C., Vidal, R., Castillo, A., Parra, D., Movil, O., Rumbos, R., Subero, L., Navarro, R., Sánchez, P., y Marín, C. (2009). Final Progress Report "Cocoa Productivity and Quality Improvement: a Participatory Approach" (2004-2009). Instituto Nacional de Investigaciones Agrícolas (INIA). Estado Miranda, Venezuela, p 1.
González, R.V., Giron, C., Sanchez, P., Castillo, A., Movil, O., Parra, D., y Vidal, R. (2011). Selection of new varieties on-farm and on-station in Venezuela. Collaborative and Participatory Approaches to Cocoa Variety Improvement. Final report of the CFC/ICCO/Bioversity project on "Cocoa Productivity and Quality Improvement: a Participatory Approach" (2004-2010). CFC, Amsterdam, The Netherlands/ICCO, London, UK/Bioversity International, Rome, Italy: Eskes AB, p 1.
Gu, Y., Yu, S., Lambert, J.D. (2014). Dietary cocoa ameliorates obesity-related inflammation in high fat-fed mice. European Journal of Nutrition,53, 149-158.
Guehi, S.T., Dabonne, S., Ban-Koffi, L., Kedjebo, D., K., y Zahouli, G.I.B. (2010). Effect of Turning Beans and Fermentation Method on the Acidity and Physical Quality of Raw Cocoa Beans. Advance Journal of Food Science and Technology, 2(3), 163-171.
Guerrero, L., Margalef, M., Pons, Z., Quiñones, M., Arola, Ll., Arola-Arnal, A., y Muguerza, B. (2013). Serum metabolites of proanthocyanidin-administered rats decrease lipid synthesis in HepG2 cells. Journal of Nutritional Biochemistry, 24, 2092-2099.
Horie, H., y Kobata, K. (2002). Major components in tea leaves. In: Muramatu et al. (eds) Health science of tea (Japanese), Gakkaishuppan center, Tokyo, pp. 26-31.
InfoGen. (2009). Grupo InfoStat, FCA, Universidad Nacional de Córdoba, Argentina.
Ishida, T., Kurata, T., Okada, K., y Wada, T. (2008). A genetic regulatory network in the development of trichomes and root hairs. Annual Review of Plant Biology,59, 365-386.
Islam, N., Lonsdale, M., Upadhyaya, N.M., Higgins, T.J., Hirano, H., y Akhurst, R. (2004). Protein extraction from mature rice leaves for two-dimensional gel electrophoresis and its application in proteome analysis. Proteomics,4, 1903-1908.
Kubo, I., y Matsumoto, A. (1984). Mollusciscides from olive Olea europaea and their efficient isolation by countercurrent chromatographies. Journal of Agricultural and Food Chemistry,32, 687-688.
Li, X., Liu, Z., Chen, Y., Wang, L.J., Zheng, Y.N., Sun, G.Z., y Ruan, C.C. (2009). Rubiacordone A: A new anthraquinone glycoside from the Roots of Rubia cordifolia. Molecules,14, 566- 572.
Lindow, S.E., y Brandl, M.T. (2003). Microbiology of the phyllosphere. Apply Environment Microbioly,69, 1875-1883.
Martinez-Esteso, M.J., Selles-Marchart, S., Vera-Urbina, J.C., Pedreño, M.A., y Bru-Martinez, R. (2009). Changes of defense proteins in the extracellular proteome of grapevine (Vitis vinifera cv. Gamay) cell cultures in response to elicitors. Journal of Proteomics,73, 331-341.
Matt, P., Krapp, A., Haake, V., Mock, H.P., y Stitt, M. (2002). Decreased Rubisco activity leads to dramatic changes of nitrate metabolism, amino acid metabolism and the levels of phenylpropanoids and nicotine in tobacco antisense RBCS transformants. The Plant Journal, 30(6), 663-677.
McDowell, E.T., Kapteyn, J., Schmidt, A., Li, Ch., Kang, J.H., Descour, A., Shi, F., Larson, M., Schilmiller, A., An, L., Jones, A.D., Pichersky, E., Soderlund, C.A., y Gang, D.R. (2011). Comparative Functional Genomic Analysis of Solanum Glandular Trichome Types. Plant Physiology, 155, 524-539.
Mishra, B.B., Kishore, N., Tiwari, V.K., Singh, D.D., y Tripathi, V. (2010). A novel antifungal anthraquinone from seeds of Aegle marmelos Correa (family Rutaceae). Fitoterapia,81, 104-107.
Morillo, F., Parra, D., Perozo, J., Movil, O., Pereira, Y., Rengifo, A., Solórzano, F., González, R., y Rengifo, J. (2012). Ataque de plantas de cacao (Theobroma cacao L.), por coquitos escopeteros (Coleoptera: Curculionidae: Scolytinae) entre 2010-2011, Estado Miranda. Instituto nacional de Investigaciones Agrícolas (INIA).Estado Miranda, Venezuela: Revista Observador del Conocimiento, p 13.
Mulekar, J.J., y Huq, E. (2014). Expanding roles of protein kinase CK2 in regulating plant growth and development. Journal of Experimental Botany,65(11), 2883-93.
Neilson, K.A. Gammulla, C.G., Mirzaei, M., Imin, N., y Haynes, P.A. (2010). Proteomic analysis of temperature stress in plants. Proteomics,10, 828-845.
Nicholson, R.L., y Hammerschmidt, R. (1992). Phenolic compounds and their role in disease resistance. Annual Review of Phytopathology,30, 369-389.
Niemenak, N., Rohsius, Ch., Elwers, S., Ndoumou, D.O., y Lieberei, R. (2006). Comparative study of different cocoa (Theobroma cacao L.) clones in terms of their phenolics and anthocianins contents. Journal of Food Composition and Analysis,19, 612-619.
Oliveira da Conceição, H.E., Mazzafera, P., Gomes da Rocha, O., y Benchimol, R.L. (1997). Biochemical Composition of Theobroma grandiflorum leaves infected by Crinipellis perniciosa. Revista Brasileira de Fisiologia Vegetal,9(2), 135-138.
Pandey, R., y Mishra, A. (2010). Antibacterial activities of crude extract of Aloe barbadensis to clinically isolated bacterial pathogens. Apply Biochemistry and Biotechnology,160, 1356-1361.
Peter, A.J., y Shanower, T.G. (1998). Plant glandular tricomes. Chemical factories with many potential uses. Resonance, March, 41-45.
Petti, S., y Scully, C. (2009). Polyphenols, oral health and disease: A review. Journal of Dentistry,37, 413-423.
Pirovani, C.P., Carvalho, H.A.S., Machado, R.C.R., Gomes, D.S., Alvim, F.C., Pomella, A.W.V., Gramacho, K.P., Cascardo, J.C.M., Amarante, G., Pereira, G., y Micheli, F. (2008). Protein extraction for proteome analysis from cacao leaves and meristems, organs infected by Moniliophthora perniciosa, the causal agent of the witches' broom disease. Electrophoresis,29, 2391-2401.
Reyes, H.E., Perez, A.Z., Reyes, L.C.; y Wagner, O.M. (1973). Catálogo de cultivares de cacao (Theobroma cacao L.). Estación Experimental de Caucagua, Estado Miranda, Venezuela, p 131.
Rusconi, M., y Conti, A. (2010). Theobroma cacao L., the food of the gods: A scientific approach beyond myths and claims. Pharmacological Research,61, 5-13.
Sarria, B., Martínez-López, S., Sierra-Cinos, JL., García-Diz, L., Mateos, R., y Bravo, L. (2014). Regular consumption of a cocoa product improves the cardiometabolic profile in healthy and moderately hypercholesterolaemic adults. British Journal of Nutrition,111, 122-134.
Tattini, M., Gravano, E., Pinelli, P., Mulinacci, N., y Romani, A. (2000). Flavonoids accumulate in leaves and glandular trichomes of Phillyrea latifolia exposed to excess solar radiation. ResearchNew Phytololgy, 148, 69-77.
Tomihama T., Nishi Y., y Arai K. (2006). The effects of exopolysaccharide production and flagellum on motility, biofilm formation, survival and virulence of Pseudomonas syringae pv. theae. Japanese Journal of Phytopathology,72, 3-13.
Van den Ende, W. (2014). Sugars take a central position in plant growth, development and, stress responses. A focus on apical dominance. Frontiers of Plant Science, 30, 247.
Vivot, E., Sánchez, C., Cacik, F., y Sequin, C. (2009). Tamizaje de la actividad antifúngica de extractos de especies de la flora de Entre Ríos. Revista Cubana de Farmacia,43 (4), 1-10.
Wagner, G.J. (1991). Secreting glandular trichomes: more than just hairs. Plant Physiology,96, 675-679.
Wagner, M., Laborem, G., Marin, C., Medina, G., y Rangel, L. (2003). Proline content in valencia orange crops during five cycles as a function of water content in Fluventic Haplustolls soils. Revista de la Facultad de Agronomía. LUZ,20, 119-130.
Wagner, G.J., Wang, E., y Shephard, R.W. (2004). New approaches for studying and exploiting the plant trichome. Annals of Botany,93, 3-11.
Yooyongwech, S., Cha-um, S., y Supaibulwatana, K. (2012). Proline related genes expression and physiological changes in indica rice response to water-deficit stress. Plant Omics Journal,5(6), 597-603.
Zapata, J., Hernandez, M.; Ojeda, M.; Benlloch, M.; Prast, E.; Tena, M.; Lopez, R.; y Jorrin, J. (2000). Toxic metals accumulation and total soluble phenolics in sunflower and tobacco plans. Plant Physiology and Biochemistry,38,178-80.
Referencias
Afoakwa, E. O., Jennifer, Q., Agnes, S. B., Jemmy, S. T., y Firibu, K. S. (2012a). Influence of pulp-preconditioning and fermentation on fermentative quality and appearance of Ghanaian cocoa (Theobroma cacao) beans. International Food Research Journal, 19(1), 127-133.
Afoakwa, E. O., Quao, J., Takrama, F. S., Budu, A. S., y Saalia, F. K. (2012b). Changes in total polyphenols, o-diphenols and anthocyanin concentrations during fermentation of pulp pre-conditioned cocoa (Theobroma cacao) beans. International Food Research Journal, 19 (3), 1071-1077.
Ávalos, A., y Pérez-Urria, E. (2009). Metabolismo secundario de plantas. Reduca (Biología). Serie Fisiología Vegetal, 2 (3), 119-145.
Bailey, B.A, Strem, M.D., y Wood, D. (2009). Trichoderma species form endophytic associations within Theobroma cacao trichomes. Mycological Research, 113, 1365–1376.
Balbuena, T.S., Dias, L.L.C., Martins, M.L.B., Chiquieri, T.B., Santa-Catarina, C., Floh, E.I.S., y Silveira, V. (2011). Challenges in proteome analyses of tropical plants. Brazilian Journal of Plant Physiology, 23(2), 91-104.
Bates, L.S., Waldron, R.P., y Teare, L.D. (1973). Rapid determination of free proline for water-stress studies. Plant Soil, 39, 205–207.
Bradford, M.M. (1976). A rapid and sensitive method for quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Analytical Biochemistry, 72, 248-254.
Cabrera-Soto, M.L., Salinas-Moreno, Y., Velázquez-Cardelas, G.A., y Espinosa-Trujillo, E. (2009). Content of soluble and insoluble phenols in the structures of corn grain and their relationship with physical properties. Agrociencia, 43, 827-839.
Carpentier, S.C., Witters, E., Laukens, K., Deckers, P., Swennen, R., y Panis, B. (2005). Preparation of protein extracts from recalcitrant plant tissues: An evalution of different methods for two-dimensional gel electrophoresis analysis. Proteomics, 5, 2497-2507.
Chen, G.Y., Yong, Z.H., Liao, Y., Zhang, D.Y., Chen, Y., Zhang, H .B., Chen, J., Zhu, J.G., y Xu, D.Q. (2005). Photosynthetic Acclimation in Rice Leaves to Free-air CO2 Enrichment Related to Both Ribulose-1,5-bisphosphate Carboxylation Limitation and Ribulose-1,5-bisphosphate Regeneration Limitation. Plant Cell Physiology, 46 (7), 1036–1045.
Cimato, A., Mattei, A., y Osti, M. (1990). Variation of polyphenol composition with harvesting period. Acta Horticulturae, 286, 453-456.
Dubois, M., Gilles, K.A., Hamiltion, JK., Rebers, PA., y Smith, F. (1956). Colorimetric method for determination of sugars and related substances. Analytical Chemistry, 28, 350–356.
Edwards, H.G.M., Villar, J.S.E., de Oliveira, L.F.C., y Le Hyaric, M. (2005). Analytical Raman spectroscopic study of cacao seeds and their chemical extracts. Analytica Chimica Acta, 538, 175-180.
Elwers, S., Zambrano, A., Rohsius, Ch., y Lieberei, R. (2009). Differences between the content of phenolic compounds in criollo, forastero and trinitario cocoa seed (Theobroma cacao L.).
European Food Research and Technology, 2009, 937-948.
Figueira, A., Lambert S., Carpenter, D., Pires, J., Cascardo, J., y Romanczyk, L. (1997). The similarity of cocoa flavour of fermented seeds from fingerprinted genotypes of Theobroma cacao L. from Brazil and Malaysia. Tropical Agriculture, 74(2), 132-139.
Friend, J. (1992). Lignin and associated phenolic acids in cell walls. In: Gurr, S.J., McPherson, M.J., Bowles, D.J. (eds.), Molecular plant pathology: a practical approach. Oxford: IRL Press, 2, pp. 51- 59.
Fry, S.C. (1987). Intercellular feruloylation of pectic polysaccharides. Planta, 171, 205-211.
Godard, S., Slacanin, I., Viret, O., y Gindro, K. (2009). Induction of defense mechanisms in grapevine leaves by emodin- and anthraquinone-rich plant extracts and their conferred resistance to downy mildew. Plant Physiology and Biochemistry, 47, 827-837.
González, R.V., Girón, C., Vidal, R., Castillo, A., Parra, D., Movil, O., Rumbos, R., Subero, L., Navarro, R., Sánchez, P., y Marín, C. (2009). Final Progress Report “Cocoa Productivity and Quality Improvement: a Participatory Approach” (2004-2009). Instituto Nacional de Investigaciones Agrícolas (INIA). Estado Miranda, Venezuela, p 1.
González, R.V., Giron, C., Sanchez, P., Castillo, A., Movil, O., Parra, D., y Vidal, R. (2011). Selection of new varieties on-farm and on-station in Venezuela. Collaborative and Participatory Approaches to Cocoa Variety Improvement. Final report of the CFC/ICCO/Bioversity project on “Cocoa Productivity and Quality Improvement: a Participatory Approach” (2004-2010). CFC, Amsterdam, The Netherlands/ICCO, London, UK/Bioversity International, Rome, Italy: Eskes AB, p 1.
Gu, Y., Yu, S., Lambert, J.D. (2014). Dietary cocoa ameliorates obesity-related inflammation in high fat-fed mice. European Journal of Nutrition, 53, 149–158.
Guehi, S.T., Dabonne, S., Ban-Koffi, L., Kedjebo, D., K., y Zahouli, G.I.B. (2010). Effect of Turning Beans and Fermentation Method on the Acidity and Physical Quality of Raw Cocoa Beans. Advance Journal of Food Science and Technology, 2(3), 163-171.
Guerrero, L., Margalef, M., Pons, Z., Quiñones, M., Arola, Ll., Arola-Arnal, A., y Muguerza, B. (2013). Serum metabolites of proanthocyanidin-administered rats decrease lipid synthesis in HepG2 cells. Journal of Nutritional Biochemistry, 24, 2092–2099.
Horie, H., y Kobata, K. (2002). Major components in tea leaves. In: Muramatu et al. (eds) Health science of tea (Japanese), Gakkaishuppan center, Tokyo, pp. 26–31.
InfoGen. (2009). Grupo InfoStat, FCA, Universidad Nacional de Córdoba, Argentina.
Ishida, T., Kurata, T., Okada, K., y Wada, T. (2008). A genetic regulatory network in the development of trichomes and root hairs. Annual Review of Plant Biology, 59, 365–386.
Islam, N., Lonsdale, M., Upadhyaya, N.M., Higgins, T.J., Hirano, H., y Akhurst, R. (2004). Protein extraction from mature rice leaves for two-dimensional gel electrophoresis and its application in proteome analysis. Proteomics, 4, 1903-1908.
Kubo, I., y Matsumoto, A. (1984). Mollusciscides from olive Olea europaea and their efficient isolation by countercurrent chromatographies. Journal of Agricultural and Food Chemistry, 32, 687-688.
Li, X., Liu, Z., Chen, Y., Wang, L.J., Zheng, Y.N., Sun, G.Z., y Ruan, C.C. (2009). Rubiacordone A: A new anthraquinone glycoside from the Roots of Rubia cordifolia. Molecules, 14, 566- 572.
Lindow, S.E., y Brandl, M.T. (2003). Microbiology of the phyllosphere. Apply Environment Microbioly, 69, 1875–1883.
Martinez-Esteso, M.J., Selles-Marchart, S., Vera-Urbina, J.C., Pedreño, M.A., y Bru-Martinez, R. (2009). Changes of defense proteins in the extracellular proteome of grapevine (Vitis vinifera cv. Gamay) cell cultures in response to elicitors. Journal of Proteomics, 73, 331–341.
Matt, P., Krapp, A., Haake, V., Mock, H.P., y Stitt, M. (2002). Decreased Rubisco activity leads to dramatic changes of nitrate metabolism, amino acid metabolism and the levels of phenylpropanoids and nicotine in tobacco antisense RBCS transformants. The Plant Journal, 30(6), 663-677.
McDowell, E.T., Kapteyn, J., Schmidt, A., Li, Ch., Kang, J.H., Descour, A., Shi, F., Larson, M., Schilmiller, A., An, L., Jones, A.D., Pichersky, E., Soderlund, C.A., y Gang, D.R. (2011). Comparative Functional Genomic Analysis of Solanum Glandular Trichome Types. Plant Physiology, 155, 524–539.
Mishra, B.B., Kishore, N., Tiwari, V.K., Singh, D.D., y Tripathi, V. (2010). A novel antifungal anthraquinone from seeds of Aegle marmelos Correa (family Rutaceae). Fitoterapia, 81, 104-107.
Morillo, F., Parra, D., Perozo, J., Movil, O., Pereira, Y., Rengifo, A., Solórzano, F., González, R., y Rengifo, J. (2012). Ataque de plantas de cacao (Theobroma cacao L.), por coquitos escopeteros (Coleoptera: Curculionidae: Scolytinae) entre 2010-2011, Estado Miranda. Instituto nacional de Investigaciones Agrícolas (INIA).Estado Miranda, Venezuela: Revista Observador del Conocimiento, p 13.
Mulekar, J.J., y Huq, E. (2014). Expanding roles of protein kinase CK2 in regulating plant growth and development. Journal of Experimental Botany, 65(11), 2883-93.
Neilson, K.A. Gammulla, C.G., Mirzaei, M., Imin, N., y Haynes, P.A. (2010). Proteomic analysis of temperature stress in plants. Proteomics, 10, 828-845.
Nicholson, R.L., y Hammerschmidt, R. (1992). Phenolic compounds and their role in disease resistance. Annual Review of Phytopathology, 30, 369-389.
Niemenak, N., Rohsius, Ch., Elwers, S., Ndoumou, D.O., y Lieberei, R. (2006). Comparative study of different cocoa (Theobroma cacao L.) clones in terms of their phenolics and anthocianins contents. Journal of Food Composition and Analysis, 19, 612-619.
Oliveira da Conceição, H.E., Mazzafera, P., Gomes da Rocha, O., y Benchimol, R.L. (1997). Biochemical Composition of Theobroma grandiflorum leaves infected by Crinipellis perniciosa. Revista Brasileira de Fisiologia Vegetal, 9(2), 135-138.
Pandey, R., y Mishra, A. (2010). Antibacterial activities of crude extract of Aloe barbadensis to clinically isolated bacterial pathogens. Apply Biochemistry and Biotechnology, 160, 1356-1361.
Peter, A.J., y Shanower, T.G. (1998). Plant glandular tricomes. Chemical factories with many potential uses. Resonance, March, 41-45.
Petti, S., y Scully, C. (2009). Polyphenols, oral health and disease: A review. Journal of Dentistry, 37, 413–423.
Pirovani, C.P., Carvalho, H.A.S., Machado, R.C.R., Gomes, D.S., Alvim, F.C., Pomella, A.W.V., Gramacho, K.P., Cascardo, J.C.M., Amarante, G., Pereira, G., y Micheli, F. (2008). Protein extraction for proteome analysis from cacao leaves and meristems, organs infected by Moniliophthora perniciosa, the causal agent of the witches’ broom disease. Electrophoresis, 29, 2391–2401.
Reyes, H.E., Perez, A.Z., Reyes, L.C.; y Wagner, O.M. (1973). Catálogo de cultivares de cacao (Theobroma cacao L.). Estación Experimental de Caucagua, Estado Miranda, Venezuela, p 131.
Rusconi, M., y Conti, A. (2010). Theobroma cacao L., the food of the gods: A scientific approach beyond myths and claims. Pharmacological Research, 61, 5–13.
Sarria, B., Martínez-López, S., Sierra-Cinos, JL., García-Diz, L., Mateos, R., y Bravo, L. (2014). Regular consumption of a cocoa product improves the cardiometabolic profile in healthy and moderately hypercholesterolaemic adults. British Journal of Nutrition, 111, 122–134.
Tattini, M., Gravano, E., Pinelli, P., Mulinacci, N., y Romani, A. (2000). Flavonoids accumulate in leaves and glandular trichomes of Phillyrea latifolia exposed to excess solar radiation. Research New Phytololgy, 148, 69-77.
Tomihama T., Nishi Y., y Arai K. (2006). The effects of exopolysaccharide production and flagellum on motility, biofilm formation, survival and virulence of Pseudomonas syringae pv. theae. Japanese Journal of Phytopathology, 72, 3–13.
Van den Ende, W. (2014). Sugars take a central position in plant growth, development and, stress responses. A focus on apical dominance. Frontiers of Plant Science, 30, 247.
Vivot, E., Sánchez, C., Cacik, F., y Sequin, C. (2009). Tamizaje de la actividad antifúngica de extractos de especies de la flora de Entre Ríos. Revista Cubana de Farmacia, 43 (4), 1-10.
Wagner, G.J. (1991). Secreting glandular trichomes: more than just hairs. Plant Physiology, 96, 675–679.
Wagner, M., Laborem, G., Marin, C., Medina, G., y Rangel, L. (2003). Proline content in valencia orange crops during five cycles as a function of water content in Fluventic Haplustolls soils. Revista de la Facultad de Agronomía. LUZ, 20, 119-130.
Wagner, G.J., Wang, E., y Shephard, R.W. (2004). New approaches for studying and exploiting the plant trichome. Annals of Botany, 93, 3–11.
Yooyongwech, S., Cha-um, S., y Supaibulwatana, K. (2012). Proline related genes expression and physiological changes in indica rice response to water-deficit stress. Plant Omics Journal, 5(6), 597-603.
Zapata, J., Hernandez, M.; Ojeda, M.; Benlloch, M.; Prast, E.; Tena, M.; Lopez, R.; y Jorrin, J. (2000). Toxic metals accumulation and total soluble phenolics in sunflower and tobacco plans. Plant Physiology and Biochemistry, 38,178-80.
Cómo citar
APA
ACM
ACS
ABNT
Chicago
Harvard
IEEE
MLA
Turabian
Vancouver
Descargar cita
CrossRef Cited-by
1. Pedro Gómez-Vera, Héctor Blanco-Flores, Ana Marta Francisco, Jimmy Castillo, Wilmer Tezara. (2021). Silicon dioxide nanofertilizers improve photosynthetic capacity of two Criollo cocoa clones (Theobroma cacao L.). Experimental Agriculture, 57(2), p.85. https://doi.org/10.1017/S0014479721000065.
2. Rafael Manuel de Jesús MEX-ALVAREZ, María Magali GUILLEN-MORALES, Patricia Margarita GARMA-QUEN, Lázaro Guadalupe RAMOS-GOMEZ. (2022). Hypolipidemic activity of Phaseolus vulgaris (Fabaceae) in mice. ECORFAN Journal Bolivia, , p.1. https://doi.org/10.35429/EJB.2022.17.9.1.9.
3. Jaime Fabián Vera Chang, Luis Humberto Vásquez Cortez, Kerly Estefanía Alvarado Vásquez, Frank Guillermo Intriago Flor, Dagmar Elena Fonseca Peralta, Piedad Francisca Yépez Macias, Cristhian Amable Vallejo Torres, Christian Simón Rivadeneira Barcia . (2024). Aprovechamiento de hojas de variedades de cacao (Theobroma cacao L.) Nacional, Forastero y Trinitario, con dos estadios fisiológicos foliares para la obtención de una infusión. Revista Agrotecnológica Amazónica, 4(1), p.e573. https://doi.org/10.51252/raa.v4i1.573.
Dimensions
PlumX
Visitas a la página del resumen del artículo
Descargas
Licencia
Derechos de autor 2015 Revista Colombiana de Biotecnología

Esta obra está bajo una licencia internacional Creative Commons Atribución 4.0.
Esta es una revista de acceso abierto distribuida bajo los términos de la Licencia Creative Commons Atribución 4.0 Internacional (CC BY). Se permite el uso, distribución o reproducción en otros medios, siempre que se citen el autor(es) original y la revista, de conformidad con la práctica académica aceptada. El uso, distribución o reproducción está permitido desde que cumpla con estos términos.
Todo artículo sometido a la Revista debe estar acompañado de la carta de originalidad. DESCARGAR AQUI (español) (inglés).















