



Publicado
2006-07-01
Revisión sobre el hongo Microcyclus ulei, agente causal del mal suramericano de la hoja del caucho
A review of the Mycrocylus ulei Ascomycetes fungus, causative agent of South American rubber-leaf blight
El hongo ascomycete Microcyclus ulei es el agente causal del SALB que es una de las enfermedades más importantes del árbol de caucho natural (Hevea brasiliensis) en América Latina y ha sido responsable de numerosas pérdidas económicas. Este hongo ha presentado alta variabilidad fisiológica y se sugiere su alta adaptabilidad, dentro de los mecanismos asociados a su virulencia se ha descrito la tolerancia al HCN. Se han obtenido clones de Hevea resistentes mediante mejoramiento genético, sin embargo, aun no son bien conocidos los mecanismos asociados a ésta. Un mayor conocimiento de este patógeno permitirá el desarrollo de nuevas estrategias de control así como el mayor entendimiento de los mecanismos asociados a resistencia del hospedero. Palabras clave: Microcyclus ulei, SALB, Hevea brasiliensis.
The Microcyclus ulei Ascomycetes fungus is the causal agent of south-American leaf blight (SALB), this being one of the most important diseases affecting the natural rubber tree (Hevea brasiliensis) in Latina-America and has been responsible for numerous economic losses. This fungus has presented high physiological variability, suggesting its great adaptability. HCN tolerance has been described as being one of the mechanisms associated with its virulence. Resistant Hevea clones have been obtained by genetic improvement; however, the mechanisms associated with this are still not well known. Greater knowledge of this pathogen will lead to developing new control strategies and better understanding of the mechanisms associated with host resistance.
Key words: Microcyclus ulei, SALB, Hevea brasiliensis.
|
|
||||
|
Rev. Colomb. Biotecnol. Vol. VIII N° 2 Diciembre 2006 50-59
|
||||
|
|
||||
|
Revisión sobre el hongo Microcyclus ulei, agente causal del mal suramericano de la hoja del caucho
A review of the Mycrocylus ulei Ascomycetes fungus, causative agent of South American rubber-leaf blight
Ibonne Aydee García Romero", Fabio AncízarAristizábar, Dolly Montoya Castaño""
|
||||
|
|
||||
|
RESUMEN
El hongo ascomycete Microcyclus ulei es el agente causal del SALB que es una de las enfermedades mÆs importantes del Ærbol de caucho natural (Hevea brasiliensis) en AmØrica Latina y ha sido responsable de numerosas pérdidas económicas. Este hongo ha presentado alta variabilidad fisiológica y se sugiere su alta adaptabilidad, dentro de los mecanismos asociados a su virulencia se ha descrito la tolerancia al HCN. Se han obtenido clones de Hevea resistentes mediante mejoramiento genØtico, sin embargo, aun no son bien conocidos los mecanismos asociados a ésta. Un mayor conocimiento de este patógeno permitirá el desarrollo de nuevas estrategias de control así como el mayor entendimiento de los mecanismos asociados a resistencia del hospedero.
Palabras clave: Microcyclus ulei, SALB, Hevea brasiliensis.
ABSTRACT
The Microcyclus ulei Ascomycetes fungus is the causal agent of south-American leaf blight (SALB), this being one of the most important diseases affecting the natural rubber tree (Hevea brasiliensis) in Latina-America and has been responsible for numerous economic losses. This fungus has presented high physiological variability, suggesting its great adaptability. HCN tolerance has been described as being one of the mechanisms associated with its virulence. Resistant Hevea clones have been obtained by genetic improvement; however, the mechanisms associated with this are still not well known. Greater knowledge of this pathogen will lead to developing new control strategies and better understanding of the mechanisms associated with host resistance.
Key words: Microcyclus ulei, SALB, Hevea brasiliensis.
|
||||
|
|
||||
|
INTRODUCCIÓN
La enfermedad llamada mal suramericano de la hoja del caucho o SALB por sus siglas en inglØs (South American Leaf Blight) es una de las enfermedades mÆs importantes del cultivo del caucho natural en América Latina y se constituyó durante las primeras dØcadas del siglo pasado en la causa
|
más importante que impidió la expansión del cultivo. En países como Brasil se presenta en todas las regiones donde se cultiva el Ærbol de caucho y ha sido causante de numerosas pérdidas económicas especialmente en la región de la Amazonia. (Gasparotto, et Æl.,1990, Kalil Filho & Junqueira, 1989). La enfermedad del SALB es ocasionada por el ataque del hongo ascomycete Microcyclus ulei y
|
|||
|
|
||||
|
Recibido: octubre 23 de 2006 Aceptado: noviembre 24 de 2006
|
||||
|
|
||||
|
**
|
Química farmacéutica, estudiante de doctorado en Ciencias Agropecuarias. Instituto de Biotecnología, Universidad Nacional de Colombia, correo electrónico: iagarciar@unal.edu.co
Ph. D en Ciencias Biológicas. Facultad de Ciencias, Universidad Nacional de Colombia. Ph. D en Ciencias Biológicas. Instituto de Biotecnología, Universidad Nacional de Colombia.
|
|||
|
|
||||
|
50
|
||||
|
|
||||
|
|
|||
|
EL HONGO Microcyclus ulei, AGENTE CAUSAL DEL MAL SURAMERICANO DE LA HOJA DEL CAUCHO
|
|||
|
|
|||
|
causa la caída prematura de las hojas jóvenes, en condiciones ambientales favorables puede llegar a causar la desfoliación total de los árboles (Gasparotto, et ál., 1984). El clima influencia considerablemente el desarrollo de la enfermedad, la duración de dos periodos de mojamiento foliar y dos de temperaturas iguales o superiores a 20 °C son factores climÆticos que determinan la ocurrencia de la enfermedad (Gasparotto, et ál., 1991). En viveros y en jardines clonales su alta incidencia ha determinado la reducción del crecimiento y disminución del porcentaje de plantas para ser injertadas.
En Ærboles adultos ataques sucesivos causan el debilitamiento de las plantas y como resultado una reducción en la producción de látex, incluso en clones muy susceptibles puede llegar a causar la muerte (Gasparotto, etál., 1984).
Clasificación taxonómica
La clasificación taxonómica del hongo se ha realizado con base en la forma perfecta o fase telomórfica que es la correspondiente a Microcyclus ulei. La fase anamórfica o conidial denominada F. heveae ha sido clasificada dentro del gØnero Fusicladium (Shubert, et ál., 2003). El hongo pertenece a la clase Ascomycete, orden Mycosphaerellales y familia Mycosphaerellaceae. El gØnero Microcyclus ha sido incluido en esta familia por Ericsson y Hawksworth (1993), quienes lo consideraron la contraparte estromÆtica de la familia Mycosphaerellaceae (Dothideales).
Dentro de las especies reportadas de este gØnero se encuentran alrededor de 15, correspondientes a hongos tropicales y subtropicales, que son patógenos biotróficos o necrotróficos de las hojas de sus hospederos. Un rango extenso de angiospermas estÆ asociado con el gØnero, y especies individuales estÆn restringidas a un hospedero particular(Cannon, etál., 1995).
Morfología y fisiología
El hongo Microcyclus ulei es un hongo biotrófico especializado que ataca únicamente plantas del gØnero Hevea y dentro de Øste, las especies Hevea brasiliensis, Hevea guinensis, Hevea benthamiana, Hevea spruceana, Hevea
|
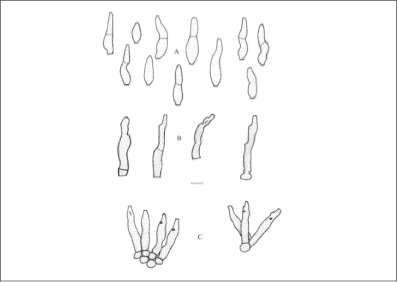
camargaona, Hevea camporum y sus híbridos. (Junqueira, et ál., 1989). Este hongo dentro de su ciclo de vida presenta tres tipos de esporas que son conidios, picnidioforos y ascosporas. La fase conidial del hongo corresponde a Fusicladium macrosporun Kuyper o Fusicladium heveae (Shubert, et ál., 2003) y es la responsable de la enfermedad en el Ærbol de caucho (Gasparotto, et ál.,1984). Los conidios se forman de manera individual, generalmente septados y a veces sin Øste, usualmente son curvados o sigmoides, lisos y hialinos (figura 1) que despuØs adquieren una coloración grisácea o verde olivo, con un tamaño aproximado de 15-65 x 6-11 um (Shubert, et ál., 2003). Las lesiones causadas por el crecimiento del hongo aparecen en las hojas, tallos, pecíolos, flores, inflorescencias y frutos jóvenes. Las manchas en las hojas se sitœan sobre la cara inferior y se encuentran esparcidas en toda la superficie foliar, son de color café grisáceo, anfígenas, variables en forma y tamaæo con un diÆmetro que oscila entre 8 mm - 10 mm y alta esporulación en clones de caucho natural susceptibles. En clones resistentes entre 1,5 y 2,0 mm con ausencia o esporulación parcial (Junqueira, etál.,1989, Araœjo, etál., 2001).
La forma telomórfica del hongo que es la denominada Microcyclus ulei, en hojas adultas forma estructuras llamadas estromas. Los estromas son de de color amarillo cafØ, globosos, generalmente adaxiales, carbonosos, superficiales amontonados y a veces formando anillos alrededor de las perforaciones de las hojas, con un tamaæo entre 200 y 400 u.m (Chee and Holliday, 1986, citado por Serie tØcnica CONIF, 1997, Schubert, et ál., 2003). Las ascas son claviformes con un tamaæo de 56-80 x 12-16 u.m, con ocho esporas, las ascosporas son alargadas septadas con un tamaæo de 18-22 x 5-8 u.m (Cannon, etál., 1995).
El hongo produce cantidades grandes de conidios los cuales son diseminados a travØs del agua lluvia o el viento siendo este œltimo el responsable de la diseminación del inoculo dentro de las mismas plantaciones así como entre plantaciones localizadas a grandes distancias (Gasparotto, et ál.; 1984). La fase picnidial conocida como Micospharella heveae no tiene importancia en la diseminación del patógeno (Chee, 1975, citado por Gasparotto, etál., 1984).
|
||
|
|
|||
|
51
|
|||
|
|
|||
|
|
|||
|
Rev. Colomb. Biotecnol. Vol. VIII N° 2 Diciembre 2006 50-59
|
|||
|
|
|||
 |
|||
|
|
|||
|
Figura 1. Fusicladium heveae. A- conidio, B - células conidiogénicas, C - conidioforos en fascículos sueltos, escala = 10 uní, tomado de (Schubert, et ál.,2003).
|
|||
|
|
|||
|
La germinación de los conidios in vitro se inicia después de tres horas de incubación. Se observa con un rÆpido crecimiento del tubo germinativo y dentro de las siguientes 24 horas las primeras ramificaciones de la hifa primaria. La germinación más efectiva ocurre a una concentración de conidios entre 1 y 5 x 105, concentraciones menores aumentan el tiempo de la germinación (Gasparotto, et ál., 1984).
Inoculaciones realizadas sobre discos de hojas de clones de Hevea resistentes y susceptibles indican que la germinación de los conidios así como la penetración miceliar entre las células de la hoja ocurren en ambos tipos de clones. La aparición de las lesiones surge al quinto día después de la inoculación y éstas sólo se presentan en hospederos susceptibles. El aumento de lesiones se observa entre el quinto y el sexto día así como la esporulación, la cantidad de esporas varía con el tipo de clon. (Hashim and Pereira, 1989). Las estructuras estromÆticas pueden llegar a aparecer después de 15 días en los clones más susceptibles (García, et ál., 1995).
|
 |
||
|
Figura 2. Foliolos en estadio C del clon PB-260 infectados con M. ulei. Tomada del archivo fotogrÆfico del Instituto de Biotecnología grupo de investigación en caucho natural.
|
|||
|
|
|||
|
52
|
|||
|
|
|||
|
|
|||
|
EL HONGO Microcyclus ulei, AGENTE CAUSAL DEL MAL SURAMERICANO DE LA HOJA DEL CAUCHO
|
|||
|
|
|||
|
Cultivo in vitro
El aislamiento del hongo se realiza a partir de material foliar infectado que presenta alto grado de esporulación, las hojas pueden estar en estado C que es el estado de maduración foliar propuesto por Halle and Martin (1968). Es conveniente realizar la siembra lo mÆs rÆpido posible con el fin de tener un mayor porcentaje de germinación de los conidios (Junqueira, etál., 1984).
Uno de los primeros reportes del cultivo in vitro del hongo fue realizado por Chee, (1978), y posteriormente por otros autores como Lieberei, et Æl., (1983), Junqueira, et Æl., (1984), y Mattos, (1999) entre otros, quienes han mejorado las metodologías de manejo in vitro del hongo propuestas inicialmente por Chee, (1978). Debido la baja tasa de crecimiento que tiene M. ulei en condiciones in vitro se han manejado diferentes tipos de medios de cultivo que van desde el uso de PDA (papa dex-trosa agar) œnicamente, hasta medios complejos como el MC4 y otros propuestos por Junqueira, et Æl., (1984).
Adicionalmente se han evaluado condiciones de fotoperiodo y temperaturas de incubación. Se han valorado tambiØn condiciones para la inducción de las diferentes fases que tiene el hongo durante su ciclo de vida. Dentro de Østas las que han presentado mayor dificultad son la obtención de micelio en corto tiempo y de conidios. Para la producción de micelio Lieberei, etál., (2006), proponen las condiciones para su obtención en cinco días en cultivo líquido de papa-sucrosa al 0.5% con concentraciones bajas de KCN, incubado a una temperatura de 23 °C en oscuridad. Para la esporulación in vitro Mattos (1999), propone la adición de suplementos como agua de coco, sin embargo los resultados obtenidos por Lieberei, et ál., (1983), muestran que la esporulación es mejor en medios poco enriquecidos como el agar papa dextrosa al 0.5% con una iluminación diaria de 90 minutos.
Variabilidad ecofisiológica
Desde la dØcada de los sesenta del siglo pasado diferentes investigadores han reportado la variabilidad fisiológica de aislamientos de M. ulei
|
obtenidos en diferentes regiones de LatinoamØrica (Chee and Holliday, 1986, citado por Serie tØcnica CONIF, 1997) y dependiendo el tipo de clones de Hevea que atacaban se hizo una clasificación inicial por razas de la 1 a la 4. Para 1966, Miller estableció una serie de clones diferenciadores para la identificación de razas fisiológicas de M. ulei. En Brasil Sudhevea (EMBRAPA), la raza 4 fue subdi-vidida en 4a, 4b y 4°. Estudios posteriores realizados por Chee, et Æl., (1986), identificaron las razas 1, 2, 3, 4, 5, 6,7 y 8, aumentando en cuatro las razas inicialmente establecidas por Miller.
Junqueira, etál., (1989), analizaron la reacción de varios clones derivados de 9 especies de caucho a 52 aislamientos de M. ulei de diferentes regiones heveícolas del Brasil y determinaron cuatro grupos distintos, de acuerdo con la esporulación de los aislamientos sobre los foliolos:
Grupo I: que corresponde a los aislamientos que esporulan en todos los clones con genes de H. Bentamiana y algunas progenies de H. brasiliensis puros.
Grupo II: aislamientos que esporulan en todas las progenies o en la mayoría de los clones de H. Brasiliensis, excepto FX985 y MDF180.
Grupo III: aislamientos que esporulan tanto en la mayoría de los clones híbridos de H. Bentamiana como en la mayoría de los clones H. brasiliensis, excepto FX985 y MDF180.
Grupo IV: que esporulan solamente en los clones de H. camporum.
Posteriormente fue descrita la alta variabilidad en aislamientos de M. ulei provenientes de diferentes regiones agroclimÆticas del Brasil, desde aislamientos recolectados en las regiones hœmedas y calientes de la región Amazónica hasta las regiones semisecas del sur del Brasil. Tambien se reportaron cambios de hospederos combinados con adaptación a un extenso rango de ambientes abióticos (Gasparotto and Junqueira, 1994). Lo anterior se ratificó con los estudios realizados por Rivano (1997), quien trabajó con 16 aislamientos de M. ulei provenientes del interior de floresta en la Guayana Francesa detectando 11 razas diferentes. El estudio hecho en la plantación de Michelin de
|
||
|
|
|||
|
53
|
|||
|
|
|||
|
|
|||
|
Rev. Colomb. Biotecnol. Vol. VIII N° 2 Diciembre 2006 50-59
|
|||
|
|
|||
|
Bahía por Mattos, etál., (2003) incluyó 50 aislamientos de M. ulei los cuales fueron evaluados en 12 clones de Hevea correspondientes a H. brasiliensis, H. pauciflora y H. benthamiana utilizando metodologías similares a las manejadas por los estudios anteriormente mencionados, pero evaluando un mayor nœmero de aislamientos. En este estudio fue posible distinguir 36 perfiles diferentes de virulencia lo cual permite suponer que se encontraron nuevas razas del patógeno. La alta variabilidad obtenida del poder patogØnico, muestra la capacidad de adaptación que tiene el hongo lo cual dificulta en un futuro la obtención de clones con resistencia durable (Mattos, et ál., 2003).
Variabilidad genØtica
Los estudios realizados para evaluar la variabilidad fisiológica de aislamientos de M. ulei de diferentes orígenes geográficos son bastantes y se ha logrado establecer un gran nœmero de razas presentes incluso en una misma plantación (Junqueira, et Æl., 1989; Rivano, 1997; Mattos, et ál., 2003), Sin embargo son pocos los estudios realizados directamente sobre el genoma del hongo como es el desarrollo de marcadores moleculares que permitan establecer si las razas hasta ahora encontradas corresponden o no a diferentes genotipos. La variabilidad genØtica se ha evidenciado con el uso de algunos marcadores moleculares como isoenzimas y mÆs recientemente con microsatØlites, no obstante estos estudios no han tenido impacto debido al nœmero pequeæo de aislamientos incluido. El estudio realizado por Junqueira, et Æl., (1987), evaluaron 7 aislamientos de M. ulei con diferentes niveles de virulencia aplicando nueve isoenzimas. Los resultados logrados permitieron encontrar que los aislamientos que difirieron en virulencia tambiØn tuvieron diferencias en los patrones isoenzimaticos obtenidos. Con los patrones de isoperoxidasas, isoestearasas e isolactato deshidrogenasas lograron diferenciar un mayor nœmero de aislados. Adicionalmente se logró establecer relación entre los patrones isoenzimÆticos de dos aislamientos con su grado de virulencia.
La detección de microsatélites específicos para M. ulei a partir de una librería enriquecida para secuencias repetidas de CA y GA, permitieron el diseæo de once juegos de primers para su amplifi-
|
cación por PCR. Éstos fueron evaluados en 11 aislamientos de M. ulei. Se encontró que 9 loci presentaron polimorfismo entre los aislados provenientes de diferentes regiones de Brasil y 5 entre los aislados provenientes de la Guayana Francesa. El locus que presentó mayor polimorfismo entre todos los aislamientos fue Mu5 (Leguen, et ál., 2006). La aplicación de esta técnica así como la de otros marcadores moleculares tipo RFLP, AFLP, ITS entre otros, en una población más grande junto con la información de características de virulencia y ecotipos permitirían un mayor entendimiento de la dinámica de las poblaciones de este patógeno.
Resistencia de Hevea al SALB
Las especies de Hevea presentan resistencia inherente al SALB lo cual ha hecho posible la selección de clones que presentan una mayor resistencia así como la obtención de nuevos clones a partir de programas de mejoramiento genØtico utilizando como principal fuente de resistencia clones de la especie Hevea benthamiana (Junqueira, etál., 1992).
La resistencia al SALB de especies de Hevea se asocia con el tamaæo de las lesiones, el periodo de latencia y la esporulación reducida del hongo en las hojas. La determinación de los periodos de latencia y la esporulación se usan comúnmente en la evaluación de resistencia horizontal en otras enfermedades. La esporulación también ha sido considerada como un test sensible para la diferenciación de razas del patógeno. En la evaluación de clones de Hevea con diferentes razas de M. ulei, el tamaño de la lesión no fue significante en tanto que la esporulación sí. Hasim, et ál., (1989), igual que Langford, sugieren tener en cuenta este œltimo parÆmetro para evaluar la resistencia horizontal en clones de Hevea y su utilidad en la selección.
Existen otros parÆmetros asociados a la resistencia de Hevea a M. ulei, entre los cuales se encuentra la producción de escopoletina que puede actuar como una barrera química, su acumulación en las hojas ha sido utilizada como un componente adicional para la descripción de la resistencia junto con los parámetros arriba descritos así como la acumulación de lignina y el número de estromas. La inoculación de 36 clones con diferentes niveles de
|
||
|
|
|||
|
54
|
|||
|
|
|||
|
|
|||
|
EL HONGO Microcyclus ulei, AGENTE CAUSAL DEL MAL SURAMERICANO DE LA HOJA DEL CAUCHO
|
|||
|
|
|||
|
resistencia, provenientes de Malasia, Brasil, Java, Guatemala, Costa de Marfil y Sri Lanka con la cepa FTP25 de M. ulei, permitió establecer que la ausencia de acumulación de escopoletina así como la de lignina en las hojas afectadas por el hongo, estuvieron relacionadas con la severidad de los síntomas de la enfermedad, sugiriendo la participación de estos compuestos en la resistencia (García, et ál., 1999). La lignificación ha sido asociada con la respuesta hipersensible, la cual es inducida rÆpidamente después de la infección, los clones totalmente resistentes presentaron una intensa lignificación, que le permitieron rodear las esporulaciones (García, et ál., 1995). Sin embargo probablemente los procesos de lignificación jueguen un papel de menor importancia que la reacción hipersensible de producción de fitoalexinas como la escopoletina, que es dominante. Es decir, las reacciones de defensa del Ærbol de caucho a M. ulei durante la infección estarían envolviendo las rutas metabólicas de síntesis de los fenilpropanoides las cuales se estarían sobreexpresando (García, et ál., 1995).
Pese a que el Hevea brasilensis es un forestal perenne se ha logrado obtener un mapa genØtico a partir de 196 individuos de la F1 producto del cruce entre un parental resistente al SALB RO38 un clon híbrido interespecífico entre H. brasiliensis x H. benthamiana y un parental susceptible de H. brasiliensis PB-260 el cual ha sido saturado mediante marcadores tipo RFLP, AFLP, microsatØlites e isoenzimas (Lespinasse, et ál., 2000).
Mediante este mapa y la evaluación de características cuantitativas asociados con resistencia al SALB como tipo de lesión (TL) y diámetro de la lesión (DL) al inocular F1 con cinco cepas de M. ulei se logró identificar seis QTL asociados con resistencia distribuidos en cinco grupos de ligamiento del mapa genØtico de RO38. Fue posible correlacionar un QTL para las cinco cepas ubicado en el cromosoma g13 cercano al marcador EM36/ 14, con las características fenotípicas DL y TL. Otros dos QTL fueron detectados para cuatro cepas uno para TL ubicado en g11 y uno ubicado en g12 para DL, los otros tres QTLs se ubicaron en los cromosomas g15, g13 y g10. Este estudio permitió la identificación de dos QTLs con un mayor efecto sobre la resistencia, mostrando que la resistencia cuantitativa es mÆs oligogØnica que poligØnica lo cual
|
es consistente con otros estudios reportados (Lespinasee, et ál., 2000). La evaluación sobre esta misma progenie de características genotípicas adicionales en condiciones naturales, permitió confirmar el mayor efecto sobre la resistencia de un QTL ubicado en g 13 cerca al marcador EM36/14, adicionalmente se estableció que este alelo denominado M13-lbn proviene desde el parental H. benthamiana del clon RO38. (Le Guen, et Æl., 2003).
Otro estudio realizado con el fin de buscar fuentes de resistencia a Microcyclus ulei y tener un mayor conocimiento de la respuesta del huØsped al ataque del patógeno, fue llevado a cabo por Hagen, et ál., (2003) en el cual se evaluó la respuesta de yuca al ser inoculada en su parte foliar con conidiosporas de M. ulei. Los resultados obtenidos muestran que la respuesta de esta planta al ataque del hongo no es una respuesta típica de resistencia no hospedero. La respuesta hipersensible en la planta producida inicialmente, es inhibida por la liberación de HCN (ácido cianúrico) de las células muertas, en tanto que el hongo sigue creciendo debido a su alta tolerancia a este compuesto. Dada la similitud de respuesta que presenta con Hevea brasiliensis se plantea la posibilidad que yuca pueda llegar a ser un nuevo huØsped del hongo biotrófico M. ulei.
La inclusión de otro tipo de estrategias moleculares enfocadas a la bœsqueda de genes asociados a la resistencia del Hevea al SALB, que impliquen menos tiempo y que complementen los resultados hasta ahora obtenidos mediante la estrategia de mapeo posicional, permitirían un apoyo más direccionado en la selección de clones así como en los programas de mejoramiento genØtico tradicionales.
Mecanismos asociados a la patogenicidad de M. ulei
Los factores de virulencia asociados a la patogenicidad de M. ulei son poco conocidos, pese ha ser un hongo ampliamente estudiado en cuanto a su morfología, fisiología y epidemiología, solamente se han reportado los estudios de Lieberei, et Æl., (1983), y Lieberei, et ál., (2006), en los que se evaluó bajo condiciones in vitro la tolerancia de M. ulei. al HCN. Este compuesto, es producido por algunas
|
||
|
|
|||
|
55
|
|||
|
|
|||
|
|
|||
|
Rev. Colomb. Biotecnol. Vol. VIII N° 2 Diciembre 2006 50-59
|
|||
|
|
|||
|
plantas como repuesta al ataque de algunos patógenos. Dentro de estas plantas cianogénicas de han reportado el genero Manihot (Hagen, et ál., 2003) y el genero Hevea (Lieberei, et ál., 1983).
El cianuro es un fuerte inhibidor de la cadena respiratoria y de la actividad de metaloenzimas, lo cual influye significativamente en el metabolismo de plantas cianogØnicas durante procesos de infección de hongos patogénicos (Lieberei, et ál., 1983). Se ha estudiado la relación de la producción de cianuro y la resistencia a patógenos en algunas plantas como el Linum usitatissimum cuando es atacado por Colletotrichum lini y en Lotus corniculatus atacado por Stephylium loti (Rissler and Millar, 1977). En el caso de Hevea la producción de HCN favorece el crecimiento de M. ulei. Lieberei, et Æl., (2006) muestran que crecimiento de M. ulei al igual que otros hongos como Neurospora crassa usan rutas metabólicas alternativas a la del citocromo para la obtención de energía, aumentando la ruta fermentativa por consiguiente la resistencia al cianuro. Adicionalmente este estudio permitió establecer el siguiente esquema del proceso de infección: durante la penetración y colonización temprana del tejido foliar del Ærbol de caucho por
|
parte de M. ulei, es liberado el HCN, a su vez el hongo adapta su metabolismo energØtico en presencia del Æcido. En contraste, la respuesta de resistencia temprana de la planta se ve disminuida por el HCN. Después de cuatro días de la colonización, se producen los conidióforos. Un día después los conidióforos rompen la epidermis disminuyendo la concentración de HCN dentro del tejido foliar lo que permite el inicio del proceso de esporulación.
Otro mecanismo muy diferente asociado a la resistencia de M. ulei al HCN es la producción de una p-glucosidasa con actividad especifica sobre los sustratos de los compuestos cianogØnicos de la planta (Lieberei, et ál., 2006), sin embargo los estudios hasta ahora realizados no son suficientes para establecer la funcionalidad y especificidad de esta enzima.
El mayor entendimiento de los factores de virulencia asociados a la patogenicidad así como la bœsqueda de genes de avirulencia que permitan el reconocimiento del patógeno por clones de Hevea permitiría contribuir al conocimiento de los mecanismos de resistencia presentes en el huØsped.
|
||
|
|
|||
 |
|||
|
|
|||
|
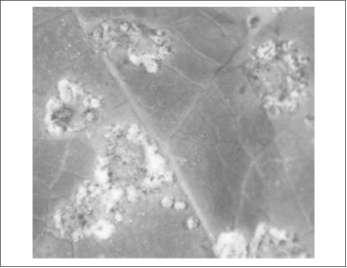
Figura 3. Crecimiento de micelio de Dicyma pulvinata sobre lesiones foliares estromÆticas causadas por M. ulei. Tomada de (Correa, et ál., 2006).
|
|||
|
|
|||
|
56
|
|||
|
|
|||
|
|
|||
|
EL HONGO Microcyclus ulei, AGENTE CAUSAL DEL MAL SURAMERICANO DE LA HOJA DEL CAUCHO
|
|||
|
|
|||
|
Control del patógeno
El SALB puede ser controlado de manera satisfactoria con el uso de fungicidas a nivel de viveros y jardines clonales. Sin embargo el uso de fungicidas en árboles adultos de plantación se hace muy limitado por la dificultad para las aplicaciones debido a la altura que alcanzan los Ærboles, e insostenible por el alto costo económico. (Nilton, et ál., 1992). Por tanto la mejor alternativa para el control del patógeno ha sido la obtención de clones resistentes a partir de programas de mejoramiento genØtico tradicionales así como el injerto de copa con clones de la especie H. benthamiana (Gasparotto, et ál., 1991).
Las aplicaciones de benomyl en jardines clonales y viveros mostraron un nivel satisfactorio para el control del hongo, presentando mayor eficiencia que fungicidas como etibendazol y mancozeb (Rocha, et Æl., 1978), Sin embargo, estudios posteriores (Junqueira, et ál., 1988) mostraron que este fungicida actúa convenientemente sólo para el control de algunos clones. La evaluación con mezclas de fungicidas como tiofanato metílico, triadimefon y methamidophos, mostró resultados similares, es decir, el control funcionó de manera diferente en los clones evaluados lo cual dependió del nivel de resistencia parcial a las diferentes razas de M. ulei. Permitiendo concluir que los clones de Hevea responden de manera diferente al control químico (Junqueira, etál., 1992).
Otra alternativa que se ha buscado mÆs recientemente es el control biológico mediante la aplicación del hongo Dicyma pulvinata que ha mostrado ser un eficiente de antagonista del M. ulei, ya que coloniza las lesiones estromÆticas (figura 3) causando la destrucción de las estructuras sexuales y en consecuencia la defoliación del árbol se disminuye. (Junqueira and Gasparotto, 1991). Se han realizado aislamientos de D pulvinata en diferentes regiones del Brasil y se ha evaluado su eficiencia de colonización de las lesiones estromáticas foliares causadas por M. ulei, de esta evaluación se seleccionaron cuatro aislamientos de 51 que muestran ser promisorios y que podrían ser utilizados como controladores biológicos del M. ulei (Correa, et ál., 2006). Sin embargo este tipo de estrategia apenas inicia sus estudios y se presenta como una solución a largo plazo.
|
Pese a que M. ulei es un hongo que se ha venido estudiando desde principios del siglo pasado y se ha avanzado en el conocimiento de su morfología, diversidad patogénica y comportamiento en campo, debido a la dificultad de su manejo in vitro aun se desconoce mucho de su fisiología y de los factores asociados a la virulencia, así como los mecanismos asociados a la resistencia por parte del gØnero Hevea.
Es relevante continuar con estudios que permitan un mayor conocimiento de este patógeno dada la importancia económica que representa el cultivo del árbol de caucho natural en países en vía de desarrollo, el cual se presenta como una alternativa económica debido a la alta demanda de caucho natural a nivel mundial. Adicionalmente el M. ulei se ha convertido en una amenaza para los países productores del Asia y África en donde actualmente no existe, ya que se ha demostrado la alta susceptibilidad que tienen los clones sembrados en sus plantaciones.
El desarrollo reciente de técnicas en biología molecular se muestra como una herramienta sólida para la generación rápida de conocimiento no solamente del hongo, también de su hospedero así como la interacción entre los dos.
AGRADECIMIENTOS
Los autores agradecen a Esperanza Torres profesora de la Facultad de Agronomía de la Universidad Nacional de Colombia y a Silvia Restrepo profesora de la Universidad de Los Andes por la revisión y correcciones hechas al manuscrito. Al proyecto "Caracterización morfológica y molecular del hongo Microcyclus ulei causante del mal suramericano de la hoja del caucho" actualmente en ejecución por la Universidad Nacional de Colombia y al Instituto Amazónico de investigaciones Científicas SINCHI con financiación del Ministerio de Agricultura.
BIBLIOGRAFÍA
Araújo, A.E.; Kalil Filho, A.; Nobrega, M.B.; Sousa, N.R.; Santos J.W.R. 2001. Reaction of ten clones of rubber (Hevea benthamiana) to three Microcyclus ulei isolates. Acta Amazónica. 31 (3): 349-356.
|
||
|
|
|||
|
57
|
|||
|
|
|||
|
|
|||
|
Rev. Colomb. Biotecnol. Vol. VIII N° 2 Diciembre 2006 50-59
|
|||
|
|
|||
|
Cannon, P.I.; Camaran C.C.; Romero A.I. 1995. Studies on biotrophic fungi from Argentina: Microcyclus porleriae, whit a key to South American species of Microcyclus. Mycol. Res 99: 353-356.
CorrŒa Marques de Mello, S.; Santos, M.; Tavares da Silva, J.B. 2006. Isolados de Dicyma pulvinata em estromas de Microcyclus ulei em seringueira. Pesq. agropec. bras.; 41(2): 359-363.
Chee, K.H.; Holliday, P. 1986. Enfermedad suramericana de la hoja del hule (caucho) Hevea. Instituto para la Investigación y Desarrollo del Hule de Malasia, MRRDB. Monografía n° 13. Presentado en la serie técnica n° 37 /Avances de la investigación en caucho natural. CONIF. 1997 Santa Fe de BogotÆ.
Chee, K.H.; Kaiming, Z.; Darmono, T.W. 1986. Ocurrence of eight races of Microcyclus ulei on Hevea in Bahía. Brasil. British Mycological Society. 87: 15-21.
Chee, K.H. 1978. South American Leaf Blight of Hevea
brasiliensis: Culture of Microcyclus ulei. Trans.
Br. Mycol. Soc. 70(3): 341. Eriksson, O.E.; Hawksworth D.L. 1993. Outline of
the ascomycetes-1993. System Ascomycetum.
12: 51-257.
García, D.; Cazaux, E.; Rivano, R; Auzac, D. 1995. Chemical and structural berriers to Microcyclus ulei, the agent of South American Leaf Blight, in Hevea spp. Eur. J. For. Path. 25: 282-292.
García, D.; TroispouxV.; Granee, N.; Rivano, R; Auzac, D. 1999. Evaluation of the resistance of 36 Hevea clones to Microcyclus ulei and relation to their capacity to accumulate scopoletin and lignins. Eur. J. For. Path. 29: 323-338.
Gasparotto, L; Lieberei, R.; Trindade, D. 1984. In vitro conidia germination of Microcyclus ulei and its sensitivity to fungicides. Fitopatología Brasileira. 9: 505-511.
Gasparotto, L.; Ferreira, F.A.; Lima, M.I.; Pereira, J.C.; Santos, A.F. 1990. Enfermedades da seringueira no Brasil. Circular TØcnica 3, EMBRAPA-CPAA, Manaus. 169 p.
Gasparotto, L.; Zambolin, L.; Junqueira, N.T.V, Mafia, L.A.; Ribeiro, F. X. 1991. Epidemiology of south American leaf blight of rubber tree. ll -Manaus región - AM. Fitopatología Brasileira. 16(1): 19-21.
Gasparotto, L.; Junqueira, N.T.V., 1994. Ecophysio-logical variability of Microcyclus ulei, causal agent of rubber tree leaf blight. Fitopatología Brasileira. 18: 22-24.
|
Gasparatto, L.; Figueredo, A.; Rezende, J.C.; Ferreira, F.A. 1997. Doencas da Seringueira no Brasil. Empresa Brasilera de Pesquisa Agropecuarias. 39-41. EMBRAPA.
Hagen, J.; Gasparotto L.; Moraes V.; Lieberei R. 2003. Reaction of cassava to Microcyclus ulei, Causal Agent of South American Leaf Blight of Rubber tree. Fitopatología Brasileira. 28(5) 477-480.
Halle, R; Martin R. 1968. Study of the growth rhythm in Hevea brasiliensis (Euphorbiaceae Cronoideae). Andansonia. 8(4): 475-503.
Hashim, I.; Pereira, J.C. 1989. Lesión Size, latent period and sporulation on leaf Discs as indicators of resistance of Hevea to Microcyclus ulei. J. nat. Rubb. Res. 4(11): 56-65.
Junqueira, N.T.V.; Chaves, G.M.; Zambolim, L.; Romero R.; Gasparotto L. 1984. Isolamento cultivo e esporulacáo agente etiologico de mal-da-folhas da seringueira M ulei. Revista Seres. 31: 322-331.
Junqueira, N.T.V; Chaves G.M.; Zambolim L.; Gasparotto L; Alienas A. C. 1986. Varibilidade fisiológica de M. ulei. Fitopatología Brasileira. 11: 823-833.
Junqueira, N.T.V.; Alfenas, A.C.; Chaves, G.M.; Zambolim, L.; Gasparotto, L. 1987. Variabilidade isoenzimática de isolados de Microcyclus ulei com diferentes niveis de virulencia. Fitopatología Brasileira. 12(3), 208-214.
Junqueira, N.T.V; Gasparotto, L.; Lieberei, R.; Normando, M.C.S.; Lima, M.I. 1989. Especialicáo fisiológica de Microcyclus ulei em diferentes especies de seringueira: Identificáo de grupos de patotipos Fitopatología Brasileira. 14(2):147. Resumo.
Junqueira, N.T.V; Gasparotto, L. 1991. Controle biológico de fungos estromáticos causadores de doencas foliares em seringueira. In: Bettiol, W. (Org.). Controle biológico de doencas de plantas. Jaguariúna, SP: EMBRAPA-CNPDA, v.1, p. 307-331. (EMBRAPA-CNPDA. Documentos 15).
Junqueira, N.T.V; Kalili Filho, A.N.; Araujo, A.E., 1992. Genética da resistencia da seringueira ao Microcyclus ulei. Fitopatología Brasileira. 14 (2): 149. Resumo
Kalil Filho, A.N.; Junqueira N.T.; 1989. Bases e procedimientos para o programa del melhoramiento de seringueira no CNPSD-Manaus, AM. Documentos 8, EMBRAPA-CNPSD, Manaus. 13 p.
Le Guen, V; Lespinasse, D.; Oliver, G; Rodier-Goud, M.; Pinard, F.; Seguin, M. 2003. Molecular
|
||
|
|
|||
|
58
|
|||
|
|
|||
|
|
|||
|
EL HONGO Microcyclus ulei, AGENTE CAUSAL DEL MAL SURAMERICANO DE LA HOJA DEL CAUCHO
|
|||
|
|
|||
|
mapping of genes conferring field resistance to South American leaf blight (Microcyclus ulei) in the rubber tree. Theor Appl Genet. 108: 160-167.
Le Guen, V.; Rodier-Goud M.; Troispoux, T.; Seguin, M. 2004. Characterization of polymorphic microsatellite markers for Microcyclus ulei, causal agent of South American Leaf blight of rubber trees. Molecular Ecology Notes. 4: 122-124.
Lespinasse, D.; Grivet, L.; Troispoux, V.; Rodier-Goud, M.; Pinard, F.; Seguin, M. 2000. Identification of QTLs involved in the resistance to South American leaf blight (Microcyclus ulei) in the rubber tree. Theor Appl Genet. 100: 975-984.
Lespinasse, D.; Rodier-Goud, M.; Grivet, L.; Leconte A,; Legnate, H.; Seguin, M. 2000. A saturated genetic linkage map of rubber tree (Hevea spp) based en RFLP, AFLP, microsatelite and isozyme markers. Theor Appl Genet. 100: 127-138.
Lieberei, R.; Schrader, A.; Biehl, B.; Chee, K. H. 1983. Effect of cyanide on Microcyclus ulei cultures. / Journal of the Rubber Research, Institute of Malaysia. 31(3): 227-235.
Lieberei, R. 2006. Physiological characteristics of Microcyclus ulei (P. Henn.) V.ARX. - A fungal pathogen of the cyanogenic host Hevea brasiliensis. Journal of Applied Botany and Food Quality 80(1): 63-68.
|
Mattos, C. 1999. Culture media with coconut water for sporulation of Microcyclus ulei. Fitopatología Brasileira. 24(3): 470.
Mattos, C.R.; García, D.; Pinard, F.; Le guen, V. 2003. Variabilidade de isolados de Microcyclus ulei no Sudeste de Bahia. Fitopatología Brasileira. 28 (5): 502-507.
Miller, J.W. 1966. Diferencial clones of Hevea for identifying races of Dothidella ulei. Plant Disease Repórter. 50:187-190.
Nilton, T.V.; Junqueira, Lima M.; Gasparotto, L.; Luis, A.J. 1992. Integrated control of rubber tree leaf Blight association between genetic resistente and chemical control. Pesq. Agropec. Bras. 27 (7): 1027-1034.
Rissler, J.F.; Millar, R.L. 1977. Contribution of a cyanide insensitive alternative respiratory system to increases in formamide hydrolase activity and to growth in Stemphyium loti. Plant Physiol. 60: 857-861.
Rivano, F. 1997. South American leaf blinght of Hevea I. Variability of Microcyclus ulei pathogenecity. Plantation Recherche, Devel. 4: 104-114.
Rocha, H.M.; Medeiros, A.G.; Vasconcelos, A.P. 1978. Comparacao de fungicidas para controle do mal-das-folhas de seringueira (Microcyclus ulei (P. Henn.) v. Arx) em viveiro. Fitopatología Brasileira. 3: 163.
Schubert, K.; Ritschel, A.; Braun, U. 2003. A monograph of Fusicladium lat. (Hyphomyce-tes). Schlenchtendalia. 9: 353-356.
|
||
|
|
|||
|
59
|
|||
|
|
|||
Cómo citar
APA
García-Romero, I. A., Ancízar Aristizábar, F. & Montoya Castaño, D. (2006). Revisión sobre el hongo Microcyclus ulei, agente causal del mal suramericano de la hoja del caucho. Revista Colombiana de Biotecnología, 8(2), 50–59. https://revistas.unal.edu.co/index.php/biotecnologia/article/view/515
ACM
[1]
García-Romero, I.A., Ancízar Aristizábar, F. y Montoya Castaño, D. 2006. Revisión sobre el hongo Microcyclus ulei, agente causal del mal suramericano de la hoja del caucho. Revista Colombiana de Biotecnología. 8, 2 (jul. 2006), 50–59.
ACS
(1)
García-Romero, I. A.; Ancízar Aristizábar, F.; Montoya Castaño, D. Revisión sobre el hongo Microcyclus ulei, agente causal del mal suramericano de la hoja del caucho. Rev. colomb. biotecnol. 2006, 8, 50-59.
ABNT
GARCÍA-ROMERO, I. A.; ANCÍZAR ARISTIZÁBAR, F.; MONTOYA CASTAÑO, D. Revisión sobre el hongo Microcyclus ulei, agente causal del mal suramericano de la hoja del caucho. Revista Colombiana de Biotecnología, [S. l.], v. 8, n. 2, p. 50–59, 2006. Disponível em: https://revistas.unal.edu.co/index.php/biotecnologia/article/view/515. Acesso em: 16 mar. 2026.
Chicago
García-Romero, Ibonne Aydee, Fabio Ancízar Aristizábar, y Dolly Montoya Castaño. 2006. «Revisión sobre el hongo Microcyclus ulei, agente causal del mal suramericano de la hoja del caucho». Revista Colombiana De Biotecnología 8 (2):50-59. https://revistas.unal.edu.co/index.php/biotecnologia/article/view/515.
Harvard
García-Romero, I. A., Ancízar Aristizábar, F. y Montoya Castaño, D. (2006) «Revisión sobre el hongo Microcyclus ulei, agente causal del mal suramericano de la hoja del caucho», Revista Colombiana de Biotecnología, 8(2), pp. 50–59. Disponible en: https://revistas.unal.edu.co/index.php/biotecnologia/article/view/515 (Accedido: 16 marzo 2026).
IEEE
[1]
I. A. García-Romero, F. Ancízar Aristizábar, y D. Montoya Castaño, «Revisión sobre el hongo Microcyclus ulei, agente causal del mal suramericano de la hoja del caucho», Rev. colomb. biotecnol., vol. 8, n.º 2, pp. 50–59, jul. 2006.
MLA
García-Romero, I. A., F. Ancízar Aristizábar, y D. Montoya Castaño. «Revisión sobre el hongo Microcyclus ulei, agente causal del mal suramericano de la hoja del caucho». Revista Colombiana de Biotecnología, vol. 8, n.º 2, julio de 2006, pp. 50-59, https://revistas.unal.edu.co/index.php/biotecnologia/article/view/515.
Turabian
García-Romero, Ibonne Aydee, Fabio Ancízar Aristizábar, y Dolly Montoya Castaño. «Revisión sobre el hongo Microcyclus ulei, agente causal del mal suramericano de la hoja del caucho». Revista Colombiana de Biotecnología 8, no. 2 (julio 1, 2006): 50–59. Accedido marzo 16, 2026. https://revistas.unal.edu.co/index.php/biotecnologia/article/view/515.
Vancouver
1.
García-Romero IA, Ancízar Aristizábar F, Montoya Castaño D. Revisión sobre el hongo Microcyclus ulei, agente causal del mal suramericano de la hoja del caucho. Rev. colomb. biotecnol. [Internet]. 1 de julio de 2006 [citado 16 de marzo de 2026];8(2):50-9. Disponible en: https://revistas.unal.edu.co/index.php/biotecnologia/article/view/515
Descargar cita
Visitas a la página del resumen del artículo
4382
Descargas
Los datos de descargas todavía no están disponibles.
Licencia
Derechos de autor 2006 Revista Colombiana de Biotecnología

Esta obra está bajo una licencia internacional Creative Commons Atribución 4.0.
Esta es una revista de acceso abierto distribuida bajo los términos de la Licencia Creative Commons Atribución 4.0 Internacional (CC BY). Se permite el uso, distribución o reproducción en otros medios, siempre que se citen el autor(es) original y la revista, de conformidad con la práctica académica aceptada. El uso, distribución o reproducción está permitido desde que cumpla con estos términos.
Todo artículo sometido a la Revista debe estar acompañado de la carta de originalidad. DESCARGAR AQUI (español) (inglés).















