



Publicado
Evaluación de diferentes combinaciones fitohormonales en la regeneración de Solanum tuberosum (Solanaceae) Var. Pastusa Suprema a partir de explantes internodales
Palabras clave:
zeatina ribósido, ácido naftalenacético, ácido gibérelico, organogénesis indirecta, zeatin riboside, naphthalene acetic acid, gibberellic acid, indirect organogenesis (es)Evaluación de diferentes combinaciones fitohormonales en la regeneración de Solanum tuberosum (Solanaceae) Var. Pastusa Suprema a partir de explantes internodales
Evaluating different phytohormone combinations in regenerating Solanum tuberosum (Solanaceae) ar. Pastusa Suprema from intermodal explants
Jenny Paola Jiménez Barreto 1 , Alejandro Chaparro-Giraldo 2, Jennifer Blanco 3,
1Bióloga, Grupo Ingeniería Genética de Plantas, Universidad Nacional de Colombia, Bogotá.2Ph. D., Profesor Asociado; director Grupo Ingeniería Genética de Plantas, Universidad Nacional de Colombia, Bogotá, Colombia. achaparrog@bt.unal.edu.co.
3Bióloga; estudiante MSc. Ciencias Agrarias; Grupo Ingeniería Genética de Plantas, Universidad Nacional de Colombia, Bogotá. jtblancom@unal.edu.co
Recibido: septiembre 15 de 2009 Aprobado: noviembre 12 de 2009
Resumen
La regeneración de plantas mediante el cultivo de tejidos es un importante componente de la biotecnología que es requerido para procesos tales como la obtención de plantas transgénicas. Se estableció un sistema eficiente de regeneración para la especie Solanum tuberosum L. var. Pastusa Suprema, susceptible de ser transformada genéticamente. Se evaluó el efecto de las fitohormonas zeatina ribósido (ZR), ácido naftalénacetico (ANA) y ácido gibérelico (AG3), utilizadas en combinaciones específicas, sobre la inducción de callo, la regeneración y el número de brotes producidos por explante. La presencia de ANA demostró ser esencial en la respuesta callogénica y regenerativa de los explantes. Se encontró que la adición de 3,0 mg/L de ZR, 0,02 mg/L de ANA y 1,0 mg/L de AG3 sobre un medio básico M-S, es una formulación hormonal adecuada para inducir el proceso de organogénesis indirecta sobre la variedad de papa Pastusa Suprema; produce callogénesis y regeneración en porcentajes superiores al 90%, con un promedio de seis regenerantes por explante.
Palabras clave: zeatina ribósido, ácido naftalenacético, ácido gibérelico, organogénesis indirecta.
Abstract
Plant regeneration using tissue culture represents an important biotechnological tool which is necessary in processes such as obtaining transgenic plants. An efficient regeneration system was established for Solanum tuberosum L. var. Pastusa Suprema; this specie is susceptible to being genetically transformed. The effect of zeatin riboside (ZR), gibberellic acid (GA3) and naphthalene acetic acid (NAA) phytohormones used in specific combinations was evaluated regarding callus induction and bud regeneration and number per explant. The presence of NAA was seen to be essential in explants callogenic and regenerative response. Adding 3.0 mg/L ZR, 0.02 mg/L NAA and 1.0mg/L GA3 to a basic M-S medium was an appropriate hormonal combination for inducing indirect organogenesis in the Pastusa Suprema potato variety; it produced more than 90% callus genesis and plant regeneration and an average of six regenerants per explant.
Key words: zeatin riboside, naphthalene acetic acid, gibberellic acid, indirect organogenesis.
Introducción
La obtención de plantas transgénicas mediada por Agrobacterium tumefasciens ha sido usada exitosamente para introducir genes foráneos en diferentes cultivares de papa (De Block, 1988; Dale y Hampson, 1995; Beaujean et al., 1998; Trujillo et al ., 2001; Sarker y Mustafa, 2002). La regeneración de plantas mediante el cultivo de tejidos es un importante componente de la biotecnología que es requerido para llevar acabo este tipo de procesos (Yasmin et al ., 2003).
La regeneración vegetal es la manifestación de la totipotencia celular, la cual puede ocurrir de manera parcial siguiendo la ruta organogénica (caulogénesis o rizogénesis) o en forma completa siguiendo la ruta embriogénica (Gómez et al .,1997). En investigaciones de transformación genética el sistema de regeneración más utilizado es la organogénesis. Mediante esta técnica se desarrollan órganos sobre partes de plantas (explantes) cultivadas in vitro, siguiendo una fase inicial de desdiferenciación, posterior a la cual tiene lugar la división celular que permite la formación de un callo, a partir del que se desarrollan órganos adventicios.
Tanto la inducción de callo como la regeneración de brotes requiere la presencia de concentraciones y combinaciones apropiadas de los reguladores de crecimiento presentes en el medio de cultivo (Yasmin et al ., 2003). Se ha demostrado que los requerimientos hormonales para que la organogénesis in vitro se lleve a cabo eficientemente son altamente dependientes del genotipo (Wheleer et al ., 1985; Carputo et al ., 1995; Dale y Hampson, 1995; Hamdi et al., 1998; Sarker y Mustafa, 2002), variando entre especies y entre variedades de una misma especie.
La relación auxina-citoquinina es de vital importancia en la organogénesis, sin embargo, algunos autores han citado la necesidad de utilizar hormonas como las gibérelinas para que la regeneración se lleve a cabo eficientemente (Webb et al ., 1983; Ochatt y Caso, 1986; Sarker y Mustafa, 2002).
En la Universidad Nacional de Colombia, Sede Bogotá, el grupo de Ingeniería Genética de Plantas (IGP), ha venido trabajando en la obtención de plantas transgénicas a partir de la variedad de papa Diacol Capiro, importante desde el punto de vista de la producción industrial de papa frita. Sin embargo, es de interés llevar a cabo este tipo de procedimientos en otras variedades de papa colombiana.
La variedad Pastusa Suprema es un cultivar obtenido mediante mejoramiento genético convencional, resultado de una labor investigativa iniciada en Nariño en 1997 en la que participaron la Universidad Nacional de Colombia, Fedepapa, el ICA y Colciencias. Fue liberada al mercado el 7 de diciembre del 2002. Esta nueva variedad presenta cualidades superiores a otros cultivares, como aptitud para procesos industriales, excelente calidad culinaria, y requerimientos significativamente menores de fungicidas. Es importante desarrollar un sistema eficiente de regeneración en esta variedad, como soporte para ulteriores estudios de transferencia de genes que permitan optimizar aún más su rendimiento.
En este trabajo se evaluó el efecto de las hormonas ácido naftalénacetico (ANA), ácido giberélico (AG3) y ZR (Zeatina Ribósido), en el comportamiento callogénico y regenerativo de Solanum tuberosum L. variedad Pastusa Suprema.
Materiales y métodos
Este trabajo se realizó como parte del proyecto Desarrollo de sistemas de ingeniería genética para variedades colombianas de papa (Solanum tuberosum), resistencia a insectos y marcadores de selección, financiado por Colciencias.
Los experimentos se iniciaron con un grupo base de plántulas in vitro (aprox. 30 plantas) de S. tuberosum Var. Pastusa Suprema, donadas por el profesor Carlos Eduardo ñustez de la Facultad de Agronomía de la Universidad Nacional de Colombia. Estas plantas fueron micropropagadas en un medio básico compuesto de sales de Murashige y Skoog (1962) y suplementado con 0,4 mg/L Tiamina, 2 mg/L ácido D-pantoténico, 0,2 mg/L ácido giberélico, 20 g/L sacarosa, 2,5 mg/L Fitagel y pH 5,7.
Los ensayos de regeneración se realizaron cuando las plantas alcanzaron 3-4 semanas de edad. Se aislaron segmentos de tallo (sin incluir las yemas) de una longitud aproximada de 5-6 mm, y se sembraron sobre medios de cultivo compuestos de sales MS y combinaciones hormonales específicas (tabla 1), realizando cambio de medio cada tres semanas. El material fue mantenido a una temperatura de 17 °C, y un fotoperiodo de 16 horas luz y 8 horas oscuridad. Los experimentos se realizaron en dos series, en una fase preeliminar se evaluaron las combinaciones correspondientes a los tratamientos 1, 2, 5, 6, 9-12 de la tabla 1, utilizando 14 explantes por tratamiento. Posteriormente, se evaluaron los tratamientos 3, 4, 7 y 8, utilizando 10 explantes por tratamiento. En todos los ensayos se realizaron cuatro repeticiones por cada tratamiento.
La eficiencia del medio de cultivo se evaluó con base a las variables: porcentaje de inducción de callos (EC/ET)*100, porcentaje de regeneración (ER/ET)*100, y número de regenerantes por explante. Donde: EC = explantes con callo, ER = explantes con regenerantes y ET = explantes totales.
Se comprobó la normalidad de los datos utilizando el test de Pearson, y se realizaron los análisis de varianza respectivos con el software estadístico SAS versión 8.0.
Resultados y discusión
Comportamiento callogénico
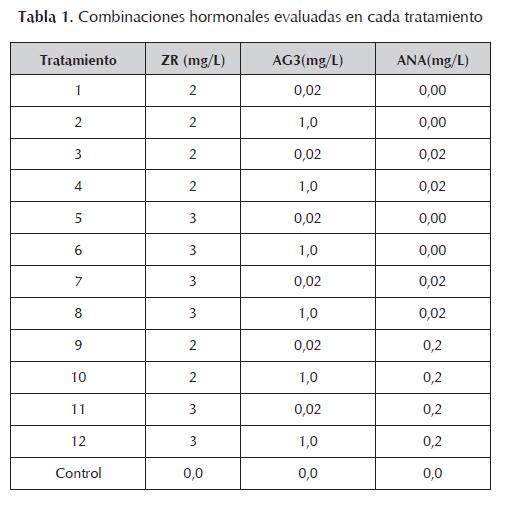
La respuesta callogénica presentada en los explantes fue diferencial según el tipo de tratamiento (figura 1). En los tratamientos en los que se utilizó la combinación de las tres hormonas (tratamientos 3, 4, 7, 8, 9, 10, 11 y 12) el proceso callogénico fue exitoso, iniciándose hacia la segunda semana, con porcentajes de inducción de callos que alcanzaron el 100%; mientras que en los tratamientos carentes de auxina (1, 2, 5 y 6) se presentó formación de callo hasta la quinta semana con un bajo porcentaje de explantes (16% ± 11,32).
La formación de callo constituye la primera fase en la regeneración mediante organogénesis indirecta. El ANA es esencial en este proceso, y su eficacia en el cultivo de tejidos ha sido ampliamente demostrada. En este trabajo se evaluaron tres niveles diferentes de la hormona: 0,0 mg/L, 0,02 mg/L y 0,2 mg/L, que influenciaron notablemente la inducción de callo.
La presencia de ANA en el medio de cultivo dio lugar a una rápida respuesta morfogénica de los explantes. Aunque algunos autores (Yasmin et al ., 2003) reportan que el aumento en la concentración de ANA da lugar a porcentajes de inducción de callo más elevados, con las concentraciones utilizadas la diferencia fue poco significativa. Sin embargo, se observaron callos más grandes en los tratamientos que contenían un nivel elevado de la hormona (figura 2), como ha sido anotado también por Turhan (2004).
En varios cultivares se ha encontrado que la auxina no es requerida para el desarrollo morfogénico del explante (Gómez et al ., 1997; Sarker y Mustafa, 2002). Para la variedad Parda Pastusa, Rodríguez et al . (2000) encontraron mejores resultados callogénicos y regenerativos en ausencia de auxina, utilizando como explante discos de hojas. Sin embargo, en los ensayos aquí descritos se encontraron resultados opuestos, lo cual indica que probablemente, además del genotipo, los requerimientos hormonales sean dependientes del tipo de explante utilizado, como ya ha sido anotado (Wheeler et al ., 1985; Ochatt y Caso, 1986; Carputo et al ., 1995; Hamdi et al ., 1998; Anjum y Ali, 2004). En este estudio también se realizaron ensayos con la variedad Parda Pastusa, pero ésta no respondió a ninguno de los tratamientos probados (datos no mostrados).
La zeatina parece promover el crecimiento de callos y la diferenciación celular (Ahloowalia, 1982); la acción conjunta de esta hormona con un nivel adecuado de auxina parece favorecer la callogénesis, como lo reveló la interacción existente entre estos dos factores.
Comportamiento regenerativo
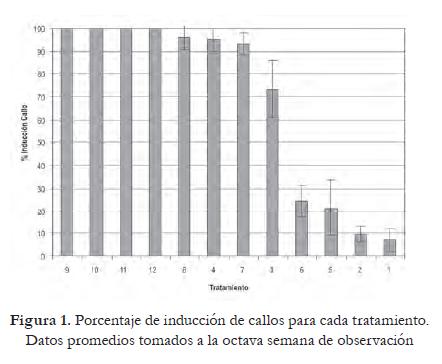
El proceso regenerativo fue influenciado significativamente por las tres hormonas, según lo demuestra el análisis estadístico (p 0,0497,<0,0001 y 0,0028 para ZR, ANA y AG3 respectivamente). Los primeros indicios de regeneración se evidenciaron hacia la tercera semana en el tratamiento 8 (ZR 3 mg/L, ANA 0,02 mg/L,AG3 1,0 mg/L), y llegaron a un valor máximo hacia la octava semana. En los tratamientos 3, 4, 7, 9-12 el proceso se inició en la cuarta semana, y en los tratamientos carentes de auxina (1, 2, 5 y 6) la regeneración de brotes inició en la quinta semana.
El efecto más marcado sobre esta variable lo tiene el nivel de auxina. Una concentración baja de ANA, como la utilizada en los tratamientos 3, 4, 7 y 8 (0,02 mg/L), demostró ser adecuada para la regeneración de brotes. Los tratamientos carentes de auxina (1, 2, 5 y 6) no alcanzaron porcentajes de regeneración apropiados (figura 3), siendo necesaria la presencia de esta hormona para que el proceso se lleve a cabo completamente. Una concentración elevada de auxina (tratamientos 9, 10, 11 y 12) parece ejercer un efecto inhibitorio sobre la formación de brotes.
La influencia de la zeatina y la zeatina ribósido en el proceso de regeneración ha sido destacada en varios trabajos. Dale y Hampsom (1995) aseguran que la adición de esta hormona en el medio acelera la formación de yemas, y Ahloowalia (1982) anota que es efectiva promoviendo la formación de brotes. El efecto de esta hormona sobre la regeneración esta estrechamente ligado al nivel de auxina presente en el medio, como lo evidencia la interacción que muestran estos dos factores en el análisis estadístico.
La presencia de ácido giberélico en el medio de cultivo estimula la iniciación y formación de tallos adventicios, así como también actúa intensificando la respuesta a otros reguladores (Webb et al ., 1983), favoreciendo la organogénesis (Ochatt y Caso, 1986). Una concentración elevada de este regulador (tratamientos 2, 4, 6, 8, 10 y 12) permitió evidenciar una mejor respuesta callogénica y regenerativa, confirmando la importancia de esta hormona en la optimización de la organogénesis in vitro.
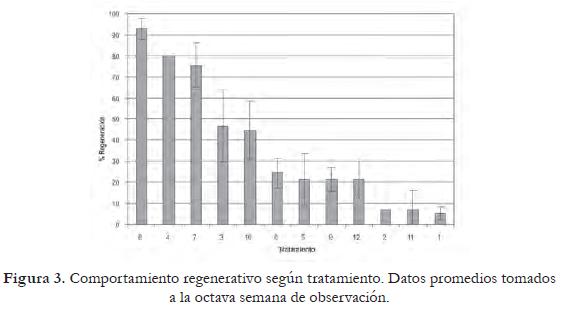
Al evaluar el efecto combinado de los tres firorreguladores (figura 4), se aprecia que el tratamiento más adecuado para el proceso de regenerativo en esta variedad es el 8 (ZR 3,0 mg/L, ANA 0,2 mg/L, AG3 1,0 mg/L).
Número de regenerantes por explante
Los datos de esta variable no se distribuyen normalmente, por esta razón no se sometieron a análisis de varianza. Sin embargo, la figura 3 permite observar que el nivel de ANA utilizado en el medio de cultivo influencia notablemente la respuesta de esta variable.
Los tratamientos carentes de auxina (1, 2, 5 y 6) presentan el mayor número de brotes por explante (7,89 ± 1,97). Los tratamientos 3, 4, 7 y 8 (ANA 0,02 mg/L), muestran un promedio de 4,69 ± 1,54 regenerantes, mientras los tratamientos que tenían altas concentraciones de ácido naftalénacetico (0,2 mg/L) presentan un número medio de regenerantes por explante notablemente menor al obtenido con un nivel bajo y en ausencia de auxina (tabla 2). Los niveles de ZR y AG3 parecen no ejercer un efecto significativo sobre este carácter.
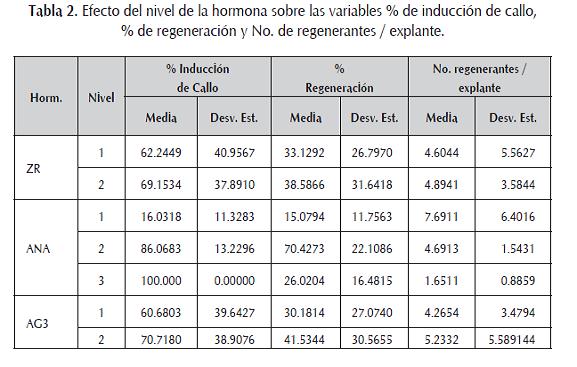
La zeatina ha demostrado aumentar el número de brotes por explante (Dale y Hampson, 1995; Park et al ., 1995; Yadav et al ., 1995), sin embargo, en las concentraciones aquí descritas no hubo una diferencia significativa, tal vez porque las dos están a un nivel elevado, siendo poco notorio el efecto de esta hormona.
Conclusiones
Los ensayos realizados demuestran la importancia de la combinación de los tres tipos hormonales en la inducción de callo y subsecuente regeneración. Probablemente, su interacción a nivel fisiológico crea las condiciones necesarias para que el tejido se desdiferencie y exprese su totipotencialidad.
Comparando los resultados obtenidos en los doce tratamientos para cada una de las variables estudiadas se puede concluir que la adición de 3,0 mg/L de zeatina ribósido, 0.02 mg/L de ácido naftalénacetico y 1,0 mg/L de ácido gibérelico en el medio de cultivo, constituye una formulación hormonal adecuada para inducir el proceso de organogénesis indirecta sobre la variedad de papa Pastusa Suprema, obteniendo porcentajes de regeneración de 92,9 % ± 4,9, y un número medio de regenerantes por explante de 6,37 ± 1,03.
Agradecimientos
Al grupo de Ingeniería Genética de Plantas de la Universidad Nacional de Colombia por su apoyo durante la realización de este trabajo, que hace parte del proyecto Desarrollo de sistemas de ingeniería genética para variedades colombianas de papa (Solanum tuberosum): marcadores de selección y resistencia a insectos, financiado por Colciencias y por la Universidad Nacional de Colombia.
Referencias bibliográficas
1 Ahloowalia, B. 1982. Plant regeneration from callus cultures in potato. Euphytica 31: 755-759.
2 Anjum, M., Ali, H. 2004. Effect of culture medium on direct organogenesis from different explants of various potato genotypes. Biotechnology 3: 187-193.
3 Anjum, M., Ali, H. 2004. Effect of culture medium on shoot initiation from calluses of different origin in Potato (Solanum tuberosum L). Biotechnology 3: 194- 199.
4 Beaujean, A., Sangwang, R., Lecardnnel, A., Sangwang- Norreel, B. 1998. Agrobacterium mediated transformation of three economically important potato cultivars using sliced internodals explants: an efficient protocol of transformation. Journal of Experimental Botany 49: 1589-1595.
5 Carputo, D., Cardi, T., Chiari, T., Ferraiolo, G. 1995. Tissue culture response in various wild and cultivated Solanum germoplasm accessions in potato breeding. Plant Cell Tiss Org Cult 41: 151-158.
6 Dale, P., Hampson, K. 1995. An assessment of morphogenic and transformation efficiency in a range of varieties of potato (Solanum tuberosum L). Euphytica 85: 101-108.
7 De Block, M. 1988. Genotype-independent leaf disc transformation of potato (Solanum tuberosum) using Agrobacterium tumefasciens. Theor Appl Genet 76: 767-774.
8 Gómez, L., Jaramillo, E., Jaramillo, S., Hoyos, R. 1997. Regeneración de plantas de papa (Solanum tuberosumL.) a partir de tejido foliar en las variedades Diacol Capiro y Parda Pastusa. Revista Papa 17: 21-25.
9 Hamdi, M., Ceballos, E., Rifer, E., Ruiz, J. 1998. Evaluación de la capacidad de regeneración de Solanum tuberosum L. Investigación Agraria y Protección Vegetales 31: 159-166.
10 Jarret, R., Hasegawa, M., Erickson, H. 1980. Effect of Medium components on shoot formation from cultured tuber disc of Potato. J Amer Soc Hort Sci 105: 238- 242.
11 Lam, S. 1977. Plantlet Formation from Tuber Disc in vitro. American Potato Journal 54: 465-468.
12 Murashige, T., Skoog, F. 1962. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol Plant 15: 473-479.
13 Ochatt, S., Caso, O. 1986. Differentials requirements among tissue source in Solanum tuberosum L. ssp. Andigena callus cultures. Turrialba 36: 363-368.
14 Park, Y., Ronis, D., Boe, A., Cheng, Z. 1995. Plant regeneration of leaf tissues of four North Dakota genotypes of Potato (Solanum tuberosum L.). American Potato Journal 72: 329-337.
15 Rodríguez, E., Trujillo, C., Orduz, S., Jaramillo, S., Hoyos, R., Arango, A. 2000. Estandarización de un medio de cultivo adecuado para la regeneración de tallos a partir de hojas utilizando dos variedades colombianas de papa (Solanum tuberosum L.). Rev Fac Nac Agr Med 53: 887-899.
16 Sarker, R., Mustafa, B. 2002. Regeneration and Agrobacterium- mediated Genetic Transformation of Two Indigenous Potato Varieties of Bangladesh. Plant Tissue Cult. 12: 69-77.
17 Trujillo, C., Rodríguez, E., Arango, S., Hoyos, R., Orduz, S., Arango, R. 2001. One-Step transformation of two Andean potato cultivars (Solanum tuberosum L. subsp. Andígena). Plant Cell Rep 20: 637-341.
18 Turhan, H. Callus induction and growth in transgenic potato genotypes. African Journal of Biotechnology 2004 (3): 375-378.
19 Yadav, N., Sticklen, M. 1995. Direct an efficient plant regeneration from leaf explants of Solanum tuberosumL. cv. Bintje. Plant Cell Reports 14: 645-647.
20 Yasmin, S., Nasirudin, K., Begum, R., Taluker, S. 2003. Regeneration and establishment of Potato plantlets through callus formation with BAP and ANA. Asian journal of plant sciences 2: 936-940.
21 Webb, K., Osifo, E., Hensahaw, G. 1983. Shoot regeneration from leaflet discs of six cultivars of potato (Solanum tuberosum subsp. Tuberosum). Plant Science Letter 30: 1-8.
22 Wheeler, V., Evans, N., Foulger, D., Webb, K., Karp, A., Franklin, J., Bright, S. 1985. Shoot formation from explant cultures of fourteen potato cultivars and studies of the cytology and morphology of regenerated plants. Ann Bot 55: 309-320.
Cómo citar
APA
ACM
ACS
ABNT
Chicago
Harvard
IEEE
MLA
Turabian
Vancouver
Descargar cita
Visitas a la página del resumen del artículo
Descargas
Licencia
Derechos de autor 2009 Revista Colombiana de Biotecnología

Esta obra está bajo una licencia internacional Creative Commons Atribución 4.0.
Esta es una revista de acceso abierto distribuida bajo los términos de la Licencia Creative Commons Atribución 4.0 Internacional (CC BY). Se permite el uso, distribución o reproducción en otros medios, siempre que se citen el autor(es) original y la revista, de conformidad con la práctica académica aceptada. El uso, distribución o reproducción está permitido desde que cumpla con estos términos.
Todo artículo sometido a la Revista debe estar acompañado de la carta de originalidad. DESCARGAR AQUI (español) (inglés).















